Série sur la nutrition osseuse non dénaturée et la santé osseuse
I. Aperçu
Nos produits nutritionnels non dénaturés pour la santé osseuse comprennent : la poudre de nutrition osseuse complète , la poudre ultra-micro de nutrition osseuse non dénaturée , la poudre ultra-micro de calcium et de protéines osseuses non dénaturées , et la poudre ultra-micro de collagène de type I non dénaturé . Ces produits sont élaborés à partir de fémurs de vache frais, adaptés à l'âge (articulations comprises), comme matière première principale, garantissant ainsi une qualité optimale de collagène, de glycosaminoglycanes et de protéines osseuses. Le procédé de fabrication repose sur une ultra-microséchage à très basse température et une technologie de séparation à très basse température par flux d'air , préservant ainsi la structure non dénaturée . Les propriétés fonctionnelles des ingrédients sont ainsi préservées, ce qui les rend idéaux pour soutenir la santé osseuse et la réparation des tissus.
La nature non dénaturée unique de ces produits leur permet de conserver leur structure active naturelle, essentielle à l'assimilation optimale des composés bioactifs par l'organisme. Cette technologie innovante assure une absorption et une efficacité supérieures pour le maintien de la santé osseuse et articulaire.
II. Proportions des composants

Le collagène est principalement composé de collagène de type I (plus de 90 %), avec une petite quantité de collagène de type II (moins de 10 %).
III. Structure active naturelle
3.1 Structure primaire
3.1.1 Collagène de type I : dominé par des séquences d’acides aminés répétitives Gly-Pro-X ou Gly-X-Hyp (liaisons peptidiques)
3.1.2 Glycosaminoglycanes
Acide hyaluronique (AH) : Une chaîne polysaccharidique linéaire simple et répétitive formée par l'alternance de liaisons glycosidiques β(1→3) et β(1→4) entre l'acide D-glucuronique et la N-acétylglucosamine.
Sulfate de chondroïtine (CS) : Une série de chaînes polysaccharidiques linéaires microscopiquement hétérogènes avec un squelette de base d'acide D-glucuronique et de N-acétylgalactosamine liés alternativement, présentant des modifications de sulfatation complexes et variables à des positions spécifiques sur la N-acétylgalactosamine (et occasionnellement sur l'acide glucuronique).
3.2 Structures supérieures
3.2.1 Collagène de type I
Structure secondaire : Chaîne peptidique α, structure hélicoïdale gauche (principalement des liaisons hydrogène)
Structure tertiaire : structure en triple hélice (liaisons peptidiques, liaisons hydrogène, forces de van der Waals, interactions hydrophobes, liaisons covalentes par condensation aldolique)
3.2.2 Structures supérieures des glycosaminoglycanes (GAG)
Structure supérieure de l'acide hyaluronique : forme en solution un réseau enroulé de manière aléatoire et fortement hydraté.
Structure supérieure du sulfate de chondroïtine : lié de manière covalente aux protéines centrales pour former des protéoglycanes.
3.3 Superstructure (Réticulation covalente supramoléculaire)
Entre les macromolécules de collagène de type I : structure de réticulation de la pyridinoline.
Entre l'ostéocalcine et l'hydroxyapatite (HAP) : Calcium chélaté par l'acide γ-carboxyglutamique.
IV. Mécanisme d'action et effets
La poudre de nutrition osseuse complète, la poudre ultrafine de nutrition osseuse non dénaturée, la poudre ultrafine de calcium et de protéines osseuses non dénaturées et la poudre ultrafine de collagène de type I non dénaturée fournissent non seulement des nutriments complets et équilibrés essentiels à la croissance et à la réparation des tissus osseux, mais offrent également les avantages des « structures actives ».
4.1 Mécanisme et effets de l'amélioration de l'absorption du calcium
4.1.1 Mécanisme : La structure supercovalente (calcium chélaté par l'acide γ-carboxyglutamique) entre l'ostéocalcine présente dans la poudre ultrafine de calcium de protéines osseuses non dénaturées et l'hydroxyapatite (HAP) est protégée par la structure tridimensionnelle de la protéine, empêchant ainsi le contact avec l'acide gastrique. Le complexe peut alors atteindre l'intestin intact. Sous l'action des enzymes digestives intestinales, il se décompose en molécules de calcium solubles, telles que le calcium chélaté par l'acide γ-carboxyglutamique ou le calcium lié à des peptides de petite taille. Ceci augmente la concentration de calcium dans l'intestin, permettant son absorption par voie paracellulaire, grâce aux gradients de concentration.
4.1.2 Effet : Cette absorption repose sur les gradients de concentration et est indépendante de la saturation de la protéine de liaison au calcium (CaBP). Chez les patients présentant une diminution du taux de CaBP liée à la croissance, au développement ou au vieillissement, entraînant une absorption limitée du calcium, ce mécanisme permet une compensation physiologique de l’absorption calcique.
4.2 Mécanisme et effet de la promotion de l'ostéogenèse calcique
4.2.1 Mécanisme : L'ostéogenèse calcique est régulée par de multiples facteurs dans les environnements sanguin et tissulaire osseux.
L'environnement sanguin est principalement influencé par le coefficient du produit calcium-phosphore. La poudre ultrafine de calcium issu de protéines osseuses non dénaturées est progressivement digérée et décomposée dans l'intestin en molécules de calcium solubles, telles que le calcium chélaté par l'acide γ-carboxyglutamique ou le calcium lié à des peptides de petite taille, permettant une absorption lente. Son rapport calcium/phosphore proche de 2:1 maximise le produit calcium-phosphore et favorise le flux de calcium du sang vers le tissu osseux.
L'environnement du tissu osseux est influencé par trois facteurs : la nano-région HAP du collagène de type I, la carboxylation des résidus d'acide glutamique dans l'ostéocalcine et la régulation par les protéines de pontage et les molécules d'adhésion osseuse.
La structure en triple hélice des fragments de collagène de type I non dénaturés qu'il contient peut activer le système lymphatique intestinal, régulant ainsi l'immunité, supprimant les réponses inflammatoires et ralentissant la dégradation du collagène de type I. Ses produits de dégradation — de petites molécules comme les peptides réticulés et les peptides conjugués — induisent la formation de la structure du collagène de type I par de multiples mécanismes (substrat, chaperon moléculaire, signalisation). Décomposés en monomères d'acides aminés, ils participent directement à la synthèse du collagène, fournissant de vastes nano-régions pour l'HAP et maximisant les nano-régions collagène-HAP.
Ses protéines osseuses non dénaturées fixent le calcium pour former de l'acide γ-carboxyglutamique lié au calcium ou de petits peptides liés au calcium. Elles peuvent également activer la γ-carboxyglutamyl carboxylase, soit comme substrats, soit par transduction de signal, catalysant la carboxylation de trois résidus d'acide glutamique sur l'ostéocalcine. Ceci crée des sites de fixation du calcium, maximisant ainsi le nombre de points d'attachement de ce dernier.
La desmine non dénaturée et l'ostéonectine non dénaturée qu'il contient peuvent favoriser la synthèse des protéines correspondantes, confiner spatialement l'HAP au sein du collagène de type I, ancrer le collagène à l'HAP et prévenir la perte de calcium osseux.
4.2.2 Effet : Le calcium peut entrer, être stocké et rester.
4.3 Mécanisme et effets de l'augmentation de la teneur en protéines osseuses
Les protéines osseuses comprennent le collagène (principalement de type I) et les glycoprotéines (notamment l'ostéocalcine, la fibronectine, l'ostéonectine et plus de dix autres). Le mécanisme d'augmentation de leur teneur est fondamentalement similaire, comme l'illustre la poudre ultrafine de collagène de type I non dénaturé.
4.3.1 Mécanisme
Mécanisme de régulation immunitaire intestinale : La poudre ultrafine de collagène de type I non dénaturé possède une structure tridimensionnelle qui résiste à la dégradation par les sucs gastriques, permettant un contact physique direct avec le système lymphatique intestinal. Ceci active les lymphocytes T immatures, induisant leur différenciation en lymphocytes T régulateurs (Tregs) spécifiques et supprimant ainsi l’attaque inappropriée du système immunitaire contre le collagène de type I. Elle favorise la sécrétion de facteurs anti-inflammatoires afin d’inhiber l’inflammation chronique et de ralentir la dégradation du collagène de type I.
Mécanisme de régulation de l'activité enzymatique intracellulaire : La poudre ultrafine de collagène de type I non dénaturée contient des structures de réticulation covalentes primaires, élevées et super-covalentes intactes. Les enzymes digestives intestinales décomposent sélectivement les réticulations covalentes primaires (liaisons peptidiques) tout en préservant les réticulations élevées et super-covalentes. Ce processus libère de petites molécules telles que des peptides réticulés et des peptides conjugués, ou ces molécules peuvent servir de substrats, de chaperons moléculaires ou de molécules de signalisation pour induire la formation de la structure du collagène de type I.
Apport de nutriments : Les peptides réticulés, les peptides conjugués et d'autres petites molécules sont ensuite décomposés en acides aminés individuels, participant directement à la synthèse du collagène.
4.3.2 Effets
Cette régulation à double action — inhibant la dégradation tout en favorisant la synthèse — offre une efficacité nettement supérieure au rôle d'apport de nutriments des protéines ou des peptides dénaturés.
4.4 Mécanisme et effets de l'amélioration des glycosaminoglycanes (GAG)
4.4.1 Mécanisme
Les particules ultrafines de glycosaminoglycanes non dénaturés libèrent des fragments d'HA, de CS et de KS. Ces fragments sont rapidement digérés en unités de faible poids moléculaire, en oligosaccharides et en disaccharidoses. Ils activent les chondrocytes, inhibent les enzymes de dégradation de la matrice (MMP ou ADAMTS), régulent l'immunité et exercent des effets anti-inflammatoires et analgésiques. À l'intérieur des cellules, ils se décomposent en molécules de glycosaminoglycanes individuelles, participant directement à la synthèse d'HA, de CS et de KS.
4.4.2 Effets
Augmente la teneur en GAG.
V. Dérivés de tissus
5.1 Déterminé par la structure des tissus
Les principales fonctions de la nutrition non dénaturée (immunomodulation, régulation de l'activité enzymatique) sont assurées par sa structure active naturelle. Cette structure présente une spécificité élevée, déterminant les propriétés tissulaires.
5.2 Nature tissulaire de la série de nutrition osseuse non dénaturée
Les poudres nutritionnelles pour os entiers, les poudres ultrafines de nutrition osseuse non dénaturées, les poudres ultrafines de calcium et de protéines osseuses non dénaturées et les poudres ultrafines de collagène de type I non dénaturé utilisent toutes des os de jarret de bovins frais et adaptés à l'âge (articulations comprises) comme matières premières. Leur protéine principale est le collagène de type I, et leur origine tissulaire doit être déterminée en fonction de ce collagène. Bien que le collagène de type I du tissu cutané partage une homologie génétique avec le tissu osseux et des séquences d'acides aminés identiques, des différences dans les modifications microenvironnementales entraînent des arrangements ultrastructuraux et d'ordre supérieur distincts. Par conséquent, le tissu cutané n'est pas considéré comme homologue à la gamme de produits nutritionnels pour os non dénaturés. Les principaux tissus homologues comprennent : le tissu osseux dur, les disques intervertébraux, les ménisques et de faibles quantités de cartilage articulaire.
VI. Population appropriée
6.1 Besoins nutritionnels : Convient à toutes les personnes présentant une carence en calcium, en particulier les nourrissons, les jeunes enfants et les personnes âgées.
6.2 Besoins de croissance osseuse : Convient aux personnes nécessitant une augmentation de la taille pendant les stades de développement et à celles cherchant à ralentir la perte de taille pendant le vieillissement.
6.2 Besoins en matière d'augmentation de la masse osseuse
6.2.1 Personnes présentant des symptômes de rachitisme pendant la croissance et le développement : tels que fermeture tardive de la fontanelle, fontanelle carrée, tête plate, poitrine en pigeon, poitrine en entonnoir, changements perlés, jambes arquées ou genoux cagneux, scoliose, etc.
6.2.2 Personnes d’âge moyen et personnes âgées : celles présentant des signes d’hyperplasie osseuse, de perte de taille, de cyphose, d’ostéoporose, de hernie discale, etc.
6.3 Besoins de réparation de la dégénérescence articulaire : Patients atteints d'arthrose dégénérative, en particulier ceux souffrant de lésions ou d'inflammation du ménisque.
6.4 Besoins en matière de guérison des lésions osseuses : Guérison des lésions osseuses ou croissance de l'implantation de ciment osseux, etc.
Nutrition articulaire non dégénérative - Série Santé articulaire
I. Aperçu
La gamme de produits nutritionnels non dénaturés Joint Health Series comprend : une poudre pour la nutrition articulaire complète, une poudre ultrafine pour la nutrition articulaire non dénaturée, une poudre ultrafine pour la nutrition du cartilage non dénaturé et une poudre ultrafine de collagène de type II non dénaturé. Tous ces ingrédients fonctionnels sont issus d'articulations ou de cartilage de genou bovins frais, adaptés à l'âge. Leur fabrication est conforme aux normes en vigueur et utilise un procédé de traitement ultrafin à très basse température (technologie brevetée) ainsi qu'une technologie de séparation à très basse température par ventilation forcée (technologie exclusive).
II. Proportions des composants

*Le collagène est principalement composé de collagène de type II (plus de 90 %), avec une petite quantité de collagène de type I (moins de 10 %).
III. Structure active naturelle
3.1 Structure primaire
3.1.1 Collagène de type II : Principalement composé de séquences d’acides aminés répétitives Gly-Pro-X ou Gly-X-Hyp (liaison peptidique)
3.1.2 Glycosaminoglycanes
Acide hyaluronique (AH) : Une chaîne polysaccharidique linéaire simple et répétitive formée par l'alternance de liaisons glycosidiques β(1→3) et β(1→4) entre l'acide D-glucuronique et la N-acétylglucosamine.
Sulfate de chondroïtine (CS) : Une série de chaînes polysaccharidiques linéaires microscopiquement hétérogènes formées par l'alternance d'unités d'acide D-glucuronique et de N-acétylgalactosamine comme squelette de base, avec des modifications de sulfatation complexes et variables se produisant à des positions spécifiques sur les unités de N-acétylgalactosamine (et occasionnellement d'acide glucuronique).
3.2 Structures supérieures
3.2.1 Collagène de type II
Structure secondaire : Chaîne peptidique α, structure hélicoïdale gauche (principalement des liaisons hydrogène)
Structure tertiaire : structure en triple hélice (liaisons peptidiques, liaisons hydrogène, forces de van der Waals, interactions hydrophobes, liaisons covalentes par condensation aldolique)
3.2.2 Glycosaminoglycanes (GAG) Structure d'ordre supérieur
Structure d'ordre supérieur de l'acide hyaluronique : forme en solution une structure de réseau enroulée de manière aléatoire et fortement hydratée.
Structure d'ordre supérieur du sulfate de chondroïtine : lié de manière covalente aux protéines centrales pour former des protéoglycanes.
3.3 Structure supramoléculaire (réticulation covalente supramoléculaire)
Réticulation intermoléculaire du collagène de type II : Hydroxypyridoline.
Ostéocalcine et hydroxyapatite (HAP) : l'acide γ-carboxyglutamique chélate le calcium.
IV. Mécanisme d'action et effets
La poudre nutritionnelle pour articulations complètes, la poudre ultrafine de nutrition articulaire non dénaturée, la poudre ultrafine de nutrition cartilagineuse non dénaturée et la poudre ultrafine de collagène de type II non dénaturée fournissent non seulement des nutriments complets et équilibrés pour les tissus articulaires, mais exercent également des effets « structurels actifs ».
4.1 Mécanisme et effets de la promotion de l'absorption du calcium
4.1.1 Mécanisme : La structure supercovalente (calcium chélaté par l’acide γ-carboxyglutamique) entre l’ostéocalcine et l’hydroxyapatite (HAP) dans la poudre ultrafine de calcium protéique osseux non dénaturé empêche le contact avec les sucs gastriques grâce à la protection de la structure tridimensionnelle de la protéine. Ceci lui permet d’atteindre l’intestin intact, où il est décomposé par les enzymes digestives intestinales en molécules de calcium solubles telles que le calcium chélaté par l’acide γ-carboxyglutamique ou le calcium lié à des peptides de petite taille, augmentant ainsi la concentration intestinale de calcium. Sous l’effet du gradient de concentration, le calcium est absorbé par voie paracellulaire.
4.1.2 Effet : Cette absorption repose sur les gradients de concentration et est indépendante de la saturation de la protéine de liaison au calcium (CaBP). Chez les patients présentant une diminution du taux de CaBP liée à la croissance, au développement ou au vieillissement, entraînant une absorption limitée du calcium, ce mécanisme permet une compensation physiologique de l’absorption calcique.
4.2 Mécanisme et effet de la promotion de l'ostéogenèse calcique
4.2.1 Mécanisme : L'ostéogenèse calcique est régulée par de multiples facteurs dans les environnements sanguin et tissulaire osseux.
L'environnement sanguin est principalement influencé par le coefficient du produit calcium-phosphore. La poudre ultrafine de calcium issu de protéines osseuses non dénaturées est progressivement digérée et décomposée dans l'intestin en molécules de calcium solubles, telles que le calcium chélaté par l'acide γ-carboxyglutamique ou le calcium lié à des peptides de petite taille, permettant une absorption lente. Son rapport calcium/phosphore proche de 2:1 maximise le produit calcium-phosphore et favorise le passage du calcium du sang vers le tissu osseux.
L'environnement du tissu osseux est influencé par trois facteurs : la nanorégion HAP du collagène de type I, la carboxylation des résidus d'acide glutamique dans l'ostéocalcine et la régulation par la bridgine et l'ostéonectine.
Les fragments de collagène de type I non dénaturés qu'il contient activent le système lymphatique intestinal, modulant l'immunité, supprimant les réponses inflammatoires et ralentissant la dégradation du collagène de type I. Ses produits de dégradation — de petites molécules comme les peptides réticulés et les peptides conjugués — agissent comme substrats, chaperons moléculaires et voies de signalisation pour induire la formation de la structure du collagène de type I. La dégradation ultérieure en monomères d'acides aminés participe directement à la synthèse du collagène, fournissant de vastes nano-régions pour l'HAP et maximisant ainsi les nano-régions de collagène-HAP.
Ses protéines osseuses non dénaturées fixent le calcium pour former de l'acide γ-carboxyglutamique lié au calcium ou de petits peptides conjugués au calcium. Elles peuvent également activer la γ-carboxyglutamique carboxylase, soit comme substrats, soit par transduction de signal, catalysant la carboxylation de trois résidus d'acide glutamique sur l'ostéocalcine. Ceci crée des sites de liaison au calcium, maximisant ainsi les points de fixation de ce dernier.
La desmine non dénaturée et l'ostéonectine non dénaturée qu'il contient peuvent favoriser la synthèse des protéines correspondantes, confiner spatialement l'HAP au sein du collagène de type I, ancrer le collagène à l'HAP et prévenir la perte de calcium osseux.
4.2.2 Effet : Le calcium pénètre, est retenu et reste.
4.3 Mécanisme et effets de la réparation du tissu cartilagineux
Le cartilage articulaire est un cartilage hyalin, dont la protéine principale est le collagène de type II.
4.3.1 Mécanisme
Mécanisme de régulation immunitaire intestinale : La poudre ultrafine de collagène de type II non dénaturé possède une structure tridimensionnelle qui résiste à la dégradation de sa conformation active naturelle par les sucs gastriques. Elle atteint l’intestin et interagit physiquement et spécifiquement avec le système lymphatique intestinal, activant les lymphocytes T naïfs pour qu’ils se différencient en lymphocytes T régulateurs (Tregs) spécifiques. Les Tregs modulent les lymphocytes T effecteurs afin de prévenir les attaques immunitaires erronées contre le collagène de type II, sécrètent des facteurs anti-inflammatoires pour supprimer l’inflammation chronique et ralentissent la dégradation du collagène de type II.
Mécanisme de régulation de l'activité enzymatique intra-organe : La poudre ultrafine de collagène de type II non dénaturée contient des structures de réticulation covalentes primaires, d'ordre supérieur et de super-ordre intactes. Les enzymes digestives intestinales dégradent sélectivement les réticulations covalentes primaires (liaisons peptidiques) tout en préservant les structures d'ordre supérieur et de super-ordre. Ceci génère de petites molécules telles que des peptides réticulés et des peptides conjugués, ou sert de substrats, de chaperons moléculaires ou de molécules de signalisation pour induire la formation de la structure du collagène de type II.
Action des nutriments : Les petites molécules comme les peptides réticulés et les peptides conjugués sont ensuite décomposées en acides aminés qui participent à la synthèse du collagène de type II.
4.3.2 Effets
Cette régulation à double action — inhibant la dégradation tout en favorisant la synthèse — offre une efficacité nettement supérieure aux effets induits par les protéines ou les peptides dénaturés.
4.4 Glycosaminoglycanes (GAG) Mécanisme et effets
4.4.1 Mécanisme
Les particules ultrafines de GAG non dénaturées fournissent des fragments d'HA, de CS et de KS, qui se digèrent facilement en fragments de faible poids moléculaire, en oligosaccharides et en unités disaccharidiques. Ces fragments activent les chondrocytes, inhibent les enzymes de dégradation de la matrice (MMP ou ADAMTS), régulent l'immunité et exercent des effets anti-inflammatoires et analgésiques. Ils peuvent également être dégradés intracellulairement en molécules de glycosaminoglycanes individuelles, participant directement à la synthèse d'HA, de CS et de KS.
4.4.2 Effets
Augmente la teneur en GAG.
V. Dérivés de tissus
5.1 Déterminé par la structure des tissus
Les principales fonctions de la nutrition non dénaturée (immunomodulation, régulation de l'activité enzymatique) sont assurées par sa structure active naturelle. Cette structure présente une spécificité élevée, déterminant les propriétés tissulaires.
5.2 Nature tissulaire de la gamme de nutrition articulaire non dénaturée
Les poudres nutritionnelles Whole Joint Nutrition Powder, Non Denatated Joint Nutrition Ultrafine Powder, Non Denatated Cartilage Nutrition Ultrafine Powder et Non Denatated Type II Collagen Ultrafine Powder utilisent toutes comme matières premières des articulations ou du cartilage de genou bovins frais et adaptés à l'âge. Leur protéine principale est le collagène de type II, provenant de tissus cartilagineux riches en ce type de collagène. Les principaux tissus homologues comprennent le cartilage articulaire et, en moindre quantité, l'os.
VI. Population appropriée
6.1 Besoins en matière de réparation des lésions articulaires
6.1.1 Besoins en matière de réparation de l'arthrose dégénérative
6.1.2 Besoins en matière de réparation des traumatismes articulaires
Nutrition cutanée non dénaturée - Gamme Santé de la peau
I. Aperçu
La gamme Skin Health Series, composée de produits nutritionnels non dénaturés, comprend : une poudre nutritive pour la peau entière, une poudre ultrafine nutritive non dénaturée pour la peau et une poudre ultrafine de collagène de type III non dénaturé. Tous ces produits sont élaborés à partir de peaux fraîches de bovins ou d'ânes d'âge approprié. Fabriqués selon les normes en vigueur grâce à une ultramicronisation à très basse température (technologie brevetée) et à une technologie de séparation à très basse température par ventilation (technologie exclusive), ces ingrédients sont des aliments fonctionnels non dénaturés.
II. Proportions des ingrédients

III. Structures actives naturelles
3.1 Structure primaire
3.1.1 Collagène de type I/III : séquences d’acides aminés répétitives (liaisons peptidiques) principalement Gly-Pro-X ou Gly-X-Hyp
3.1.2 Glycosaminoglycanes
Acide hyaluronique (AH) : Une chaîne polysaccharidique linéaire simple et répétitive formée par l'alternance de liaisons glycosidiques β(1→3) et β(1→4) entre l'acide D-glucuronique et la N-acétylglucosamine.
Sulfate de chondroïtine (CS) : Une série de chaînes polysaccharidiques linéaires microscopiquement hétérogènes formées par l'alternance d'unités d'acide D-glucuronique et de N-acétylgalactosamine comme squelette de base, avec des modifications de sulfatation complexes et variables se produisant à des positions spécifiques sur les unités de N-acétylgalactosamine (et occasionnellement d'acide glucuronique).
3.2 Structures supérieures
3.2.1 Collagène de type I/III
Structure secondaire : Chaîne peptidique α, structure hélicoïdale gauche (principalement des liaisons hydrogène)
Structure tertiaire : structure en triple hélice (liaisons peptidiques, liaisons hydrogène, forces de van der Waals, interactions hydrophobes, liaisons covalentes par condensation aldolique)
3.2.2 Structures supérieures des glycosaminoglycanes (GAG)
Structure supérieure de l'acide hyaluronique : forme en solution un réseau enroulé de manière aléatoire et fortement hydraté.
Structure supérieure du sulfate de chondroïtine : les protéines centrales se lient de manière covalente pour former des protéoglycanes.
3.3 Superstructure (Réticulation covalente supramoléculaire)
Réticulation intermoléculaire du collagène de type I : Hydroxylysine pyridinoline et lysine pyridinoline.
IV. Mécanisme d'action et effets
La poudre Whole Skin Nutrition, la poudre ultrafine de nutrition cutanée non dénaturée et la poudre ultrafine de collagène de type III non dénaturée fournissent non seulement des nutriments complets et équilibrés essentiels à la croissance et à la réparation des tissus cutanés, mais offrent également les effets des « structures actives ».
4.1 Mécanisme et effets de l'amélioration du collagène
4.1.1 Mécanisme
Mécanisme de régulation immunitaire intestinale : La poudre ultrafine de collagène de type I/III non dénaturé possède une structure tridimensionnelle qui résiste à la dégradation de sa structure active naturelle par les sucs gastriques. Elle atteint l’intestin et interagit physiquement avec le système lymphatique intestinal, activant les lymphocytes T naïfs pour qu’ils se différencient en lymphocytes T régulateurs (Tregs) spécifiques. Les Tregs modulent les lymphocytes T effecteurs afin de prévenir les attaques immunitaires erronées contre le collagène de type I/III non dénaturé, sécrètent des facteurs anti-inflammatoires pour supprimer l’inflammation chronique et ralentissent la dégradation de ce collagène.
Mécanisme de régulation de l'activité enzymatique intracellulaire : La poudre ultrafine de collagène I/III non dénaturée contient des structures de réticulation covalentes primaires, d'ordre élevé et super-ordre intactes. Les enzymes digestives intestinales peuvent sélectionner et décomposer spécifiquement les structures de réticulation covalentes primaires (liaisons peptidiques) tout en préservant les structures de réticulation covalentes d'ordre élevé et super-ordre. Ce processus génère de petites molécules telles que des peptides réticulés et des peptides conjugués, qui induisent ensuite la formation de structures de collagène de type I/III par de multiples voies, notamment en servant de substrats, de chaperons moléculaires ou de molécules de signalisation.
Fonction nutritive : Les petites molécules comme les peptides réticulés et les peptides conjugués peuvent se décomposer davantage en molécules d'acides aminés simples, participant directement à la synthèse du collagène.
4.1.2 Effet
Cette régulation à double action — inhibant la dégradation tout en favorisant la synthèse — offre une efficacité nettement supérieure à la fonction nutritive assurée par les protéines ou les peptides dénaturés.
4.2 Mécanisme et effet de l'amélioration des glycosaminoglycanes (GAG)
4.2.1 Mécanisme
Les particules ultrafines de glycosaminoglycanes non dénaturés fournissent des fragments d'HA et de CS qui se digèrent facilement en fragments de faible poids moléculaire, en oligosaccharides et en unités disaccharidiques. Ces fragments inhibent les enzymes de dégradation de la matrice (MMP ou ADAMTS), régulent les réponses immunitaires pour des effets anti-inflammatoires et analgésiques, et peuvent se décomposer intracellulairement en molécules de glycosaminoglycanes individuelles pour participer directement à la synthèse d'HA et de CS.
4.2.2 Effets
Améliore la teneur en GAG.
V. Propriétés dérivées des tissus
5.1 Propriétés tissulaires déterminées par la structure
Les principales fonctions de la nutrition non dénaturée (immunomodulation, régulation de l'activité enzymatique) sont assurées par sa structure naturellement active. Cette structure présente une spécificité élevée, sa composition déterminant les propriétés tissulaires.
5.2 Propriétés tissulaires de la gamme de nutrition cutanée non dénaturée
Les poudres Whole Skin Nutrition, Skin Nutrition Ultrafine et Non-Denatated Type III Collagen Ultrafine sont toutes élaborées à partir de peau fraîche d'âne ou de vache, adaptée à l'âge de l'animal. Leurs principales protéines sont le collagène de type I et III, provenant de tissus cutanés riches en ces types de collagène. Le principal tissu homologue est le tissu cutané.
VI. Populations appropriées
6.1 Soin anti-âge de la peau
6.2 Réparation des lésions cutanées
Nutrition cérébrale non dénaturée - Santé du cerveau
I. Aperçu
La poudre neurotrophique ultrafine de cerveau non dénaturé est un ingrédient alimentaire fonctionnel non dénaturé, produit à partir de cerveau frais de bovin ou de porc. Elle est fabriquée conformément aux normes en vigueur, grâce à un broyage ultrafin à très basse température (technologie brevetée d'invention nationale) et à une technologie de séparation à très basse température par ventilation mécanique (technologie exclusive).
II. Proportions des composants
2.1 Classification générale
Eau, lipides, protéines, sels inorganiques, autres molécules essentielles. Les sels inorganiques et les molécules essentielles sont présents en très faibles concentrations.

2.2 Composition protéique
Les protéines cérébrales représentent environ 30 % du poids sec et peuvent être classées selon leur fonction :
2.2.1 Protéines structurales : protéines microtubulaires, microfilaments (actine), protéines neurofilamentaires.
2.2.2 Protéines associées à la synapse : protéines de canal ionique, protéines de récepteur de neurotransmetteur, protéines associées aux vésicules, protéines d'échafaudage.
2.2.3 Protéines associées à la myéline : protéine basique de la myéline, protéines protéolipidiques, glycoprotéines associées à la myéline.
2.2.4 Protéines liées au métabolisme et à l'énergie : protéines mitochondriales, hexokinase, énolase
2.2.5 Facteurs neurotrophiques :
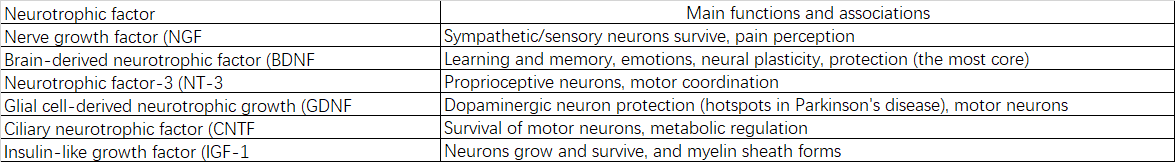
2.2.6 Protéines de signalisation : protéines G, protéines kinases, phosphatases, etc.
2.2.7 Protéines associées à la maladie : β-amyloïde, protéine tau, α-synucléine.
2.3 Composition lipidique
2.3.1 Phospholipides : cérébroside, phosphatidylinositol, phosphatidylsérine, sphingomyéline, lécithine, etc.
2.3.2 Cholestérol
2.2.3 Matières grasses neutres
Dans les phospholipides et les graisses neutres, on trouve des acides gras spécifiques : les acides gras saturés représentent environ 30 % + , tandis que les acides gras hautement insaturés représentent environ 60 % + (ARA, DHA, ALA, EPA).
III. Relation entre les protéines cérébrales et le vieillissement
Le vieillissement cérébral n'est pas un événement isolé, mais un processus systémique et progressif impliquant le déséquilibre graduel du réseau d'homéostasie protéique du cerveau, l'épuisement progressif des protéines fonctionnelles et l'accumulation progressive de protéines anormales.
3.1 Déclin global du réseau d'homéostasie protéique
Les trois principaux systèmes cellulaires assurant le contrôle de la qualité des protéines présentent tous un déclin fonctionnel :
3.1.1 Diminution de l'efficacité de la synthèse et du repliement des protéines
Les cellules vieillissantes subissent une diminution des taux et de la fidélité de la synthèse protéique, ce qui augmente la probabilité de produire des protéines mal repliées.
La réponse des chaperons moléculaires induite par le stress devient moins efficace, ne parvenant plus à aider correctement les protéines nouvellement synthétisées à se replier correctement ni à réparer les protéines endommagées.
3.1.2 Diminution de la capacité du système de dégradation des protéines
Le système ubiquitine-protéasome présente une activité réduite, incapable d'éliminer rapidement les protéines endommagées ou mal repliées ; l'assemblage du protéasome est également altéré par les dommages oxydatifs, accélérant son déclin fonctionnel.
Le système autophagie-lysosome, et plus particulièrement la macroautophagie et l'autophagie chaperonnée, présente une efficacité considérablement réduite. Ceci empêche l'élimination efficace des agrégats protéiques anormaux et des organites endommagés (par exemple, les mitochondries).
3.1.3 Altération du transport et de la distribution des protéines
Un dysfonctionnement des protéines structurales empêche l'acheminement efficace des protéines et des mitochondries nécessaires à la synapse vers l'extrémité distale, tandis que les déchets ne peuvent être rapidement transportés vers le corps cellulaire pour y être traités.
3.2 Accumulation et toxicité des protéines anormales
La défaillance du réseau d'homéostasie entraîne l'accumulation progressive de diverses « protéines déchet » dans le cerveau, endommageant directement les neurones.
3.2.1 Accumulation de protéines mal repliées et sujettes à l'agrégation
Des oligomères solubles de protéines telles que la β-amyloïde, la protéine tau et l'α-synucléine apparaissent.
3.2.2 Accumulation de protéines anormalement modifiées après la traduction
Protéines modifiées par les AGE : les produits de glycation avancée s’accumulent dans le cerveau vieillissant, provoquant une réticulation des protéines, un raidissement, une perte de fonction et l’activation des voies inflammatoires.
Protéine Tau hyperphosphorylée : la protéine Tau normale se détache des microtubules, perdant ainsi sa fonction de stabilisation du cytosquelette.
3.3 Perte de protéines fonctionnelles clés
Outre l’augmentation des « déchets », les protéines « utiles » diminuent ou deviennent également inefficaces.
3.3.1 Diminution des protéines associées aux synapses
La régulation négative de protéines présynaptiques (par exemple, la synaptophysine, SNAP-25) et postsynaptiques (par exemple, PSD-95, les récepteurs NMDA/AMPA) clés provoque directement une atrophie structurelle synaptique et un déclin fonctionnel, constituant la base moléculaire directe des troubles de l'apprentissage et de la mémoire.
3.3.2 Diminution des facteurs neurotrophiques et de leurs récepteurs
L'affaiblissement de la synthèse et de la signalisation du facteur neurotrophique dérivé du cerveau (BDNF), du facteur de croissance nerveuse (NGF) et d'autres facteurs entraîne des signaux de soutien insuffisants pour la survie, la croissance et la plasticité neuronales.
3.3.3 Diminution des protéines de réparation de l'ADN et de l'activité télomérase
Cela entraîne une instabilité génomique et une sénescence cellulaire, compromettant la survie neuronale à long terme.
3.4 Conséquences systémiques
Les altérations susmentionnées au niveau des protéines déclenchent une cascade de réactions aboutissant au vieillissement cérébral :
3.4.1 Dysfonctionnement mitochondrial : Les protéines endommagées s'accumulent dans les mitochondries, réduisant la production d'ATP et exacerbant le stress oxydatif.
3.4.2 Neuroinflammation chronique : Les cellules gliales (microglie et astrocytes) sont activées de manière persistante par des protéines anormales (par exemple, les oligomères Aβ), passant de « gardiens » à « destructeurs » en libérant des facteurs pro-inflammatoires qui endommagent les neurones sains.
3.4.3 Perte de plasticité synaptique : des déséquilibres dans les protéines clés affaiblissent la potentialisation à long terme, altérant la formation et la consolidation de la mémoire.
3.4.4 Efficacité réduite du réseau neuronal : Les dommages aux protéines de myéline dans la substance blanche et l'altération du transport axonal ralentissent la transmission de l'information entre les régions du cerveau et dégradent la synchronisation.
3.4.5 Dérégulation de l'autophagie et de l'apoptose : conduit finalement à une perte neuronale sélective, en particulier dans des régions critiques comme l'hippocampe et le cortex préfrontal.
IV. Mécanisme d'action et effets de la nutrition cérébrale non dégénérative
La nutrition cérébrale non dégénérative fournit non seulement des nutriments complets et équilibrés, essentiels à la croissance et à la réparation des tissus cérébraux (protéines, phospholipides, HA, ALA, EPA, etc.), mais surtout leurs structures actives naturelles. Ces structures possèdent des fonctions spécifiques qui dépassent le simple rôle nutritionnel.
4.1 Inhibition de la dégradation des protéines cérébrales fonctionnelles
Mécanisme d'action : La structure tridimensionnelle de la poudre ultrafine neurotrophique cérébrale non dénaturée résiste à la dégradation par les sucs gastriques, permettant un contact physique direct avec les systèmes intestinal et lymphatique. Ceci active les lymphocytes T immatures, induisant leur différenciation en lymphocytes T régulateurs (Tregs) spécifiques. Les Tregs modulent les lymphocytes T effecteurs afin de prévenir les attaques immunitaires erronées contre les protéines cérébrales, sécrètent des facteurs anti-inflammatoires pour supprimer l'inflammation chronique et ralentissent la dégradation des protéines cérébrales.
4.2 Favoriser la formation structurale et la réparation anormale des protéines cérébrales
Mécanisme : La poudre neuro-ultrafine de cerveau non dénaturée contient des structures de réticulation covalentes primaires et secondaires intactes. Les enzymes digestives intestinales décomposent sélectivement les réticulations covalentes primaires (liaisons peptidiques) tout en préservant les réticulations covalentes secondaires. Ce processus produit de petites molécules telles que des peptides réticulés et des peptides conjugués, ou sert de substrat à des enzymes spécifiques pour améliorer leur activité catalytique ; ou agit comme chaperon moléculaire pour réparer les structures anormales ; ou induit la formation de structures protéiques cérébrales et la réparation des structures anormales par de multiples voies telles que la transduction du signal.
4.3 Apport en nutriments
Mécanisme : De petites molécules comme les peptides réticulés et les peptides conjugués peuvent se décomposer davantage en acides aminés individuels, participant directement à la synthèse des protéines cérébrales.
4.4 Effet
Ce double effet régulateur, consistant à inhiber la dégradation et à promouvoir la synthèse, offre une efficacité nettement supérieure au rôle d'apport de nutriments des protéines ou des peptides dénaturés.
V. Dérivés de tissus
5.1 Régulation dépendante de la structure et dérivée des tissus
Les principales fonctions de la nutrition non dénaturée (immunomodulation, régulation de l'activité) sont assurées par des structures naturellement actives. Ces structures présentent une spécificité élevée, n'exerçant d'effets régulateurs que sur les protéines ayant des structures identiques.
5.2 Origine du tissu correspondant
La nutrition cérébrale non dénaturée utilise des tissus cérébraux frais de bovins ou de porcs, notamment le cerveau, le mésencéphale et le cervelet. Par conséquent, ses structures protéiques sont homologues à celles présentes dans le cerveau, le mésencéphale et le cervelet.
VI. Populations appropriées
6.1 Personnes nécessitant une croissance et un développement cérébral : Nourrissons, jeunes enfants et personnes ayant besoin d’un soutien au développement cérébral et neuronal.
6.2 Personnes recherchant un traitement anti-âge cérébral ou un traitement des dysfonctionnements cérébraux.
Protéines musculaires non dénaturées - Santé musculaire
I. Aperçu
La poudre ultrafine de protéines musculaires non dénaturées est un ingrédient alimentaire fonctionnel non dénaturé, produit à partir de filet de bœuf ou de blanc de poulet frais. Sa fabrication est conforme aux normes en vigueur et utilise un broyage ultrafin à très basse température (technologie brevetée au niveau national) et une technologie de séparation à très basse température basée sur le principe de l'enceinte à air (technologie exclusive).
II. Rapports et fonctions de composition
2.1 Classification générale

2.2 Composition et fonctions des protéines
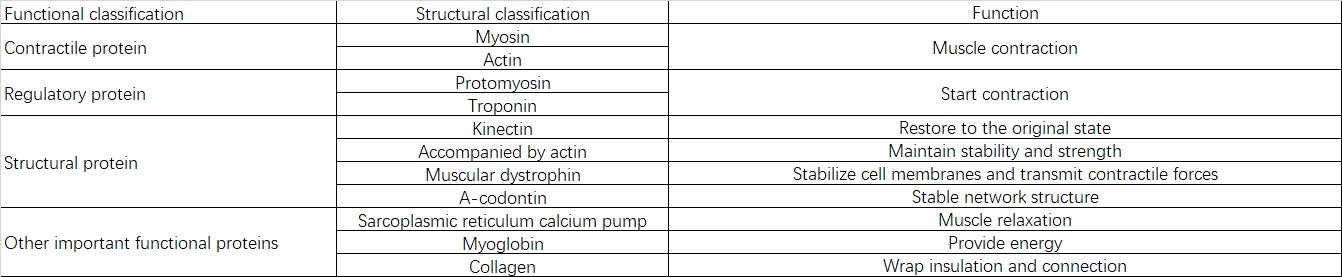
III. Causes de la sarcopénie
La sarcopénie se caractérise par une perte progressive de masse, de force et de fonction musculaire squelettique, résultant de multiples facteurs interdépendants.
3.1 Déséquilibre entre la synthèse et la dégradation des protéines (pathologie centrale)
3.1.1 Synthèse réduite
Le taux de synthèse des protéines musculaires diminue avec l'âge. Même stimulée (par exemple par l'alimentation ou l'exercice), la réponse synthétique s'affaiblit, entraînant une résistance à la synthèse protéique.
3.1.2 Dégradation accrue
Suractivation du système ubiquitine-protéasome et du système autophagie-lysosome, associée à une régulation positive de gènes tels que Atrogin-1 et MuRF1.
3.2 Dérégulation des voies hormonales et de signalisation
3.2.1 Diminution des hormones anabolisantes
Altération significative de l'axe hormone de croissance/facteur de croissance analogue à l'insuline-1.
Diminution des taux d'hormones sexuelles (testostérone/œstrogènes).
3.2.2 Inhibition des voies anaboliques :
Résistance à l'insuline : diminution de l'efficacité de l'absorption et de l'utilisation des acides aminés dans les muscles.
Suppression des principales voies anaboliques : par exemple, diminution de la capacité d’activation de la voie de signalisation mTOR.
Augmentation de la myostatine : inhibe la croissance musculaire.
Signaux cataboliques renforcés : une augmentation relative ou absolue des glucocorticoïdes favorise la dégradation des protéines.
3.3 Inflammation chronique
Le vieillissement s’accompagne d’une inflammation systémique, chronique et de bas grade avec une élévation des cytokines pro-inflammatoires (par exemple, IL-6, TNF-α).
IV. Mécanisme d'action et effets
La poudre ultrafine de protéines musculaires non dénaturées fournit non seulement des nutriments complets et équilibrés essentiels à la croissance musculaire, mais possède également des propriétés « structurelles actives ».
4.1 Mécanisme et effets de l'inhibition de la dégradation
Mécanisme d'action : La micropoudre de protéines musculaires non dénaturées contient de multiples protéines musculaires non dénaturées, conservant leur structure tridimensionnelle. Résistant à la dégradation par les sucs gastriques, ces protéines atteignent intactes l'intestin grêle. Elles sont phagocytées par les cellules M, permettant un contact physique spécifique avec le système lymphatique intestinal. Ce processus active les lymphocytes T immatures, induisant leur différenciation en lymphocytes T régulateurs (Tregs) spécifiques. Il module ainsi les attaques erronées des lymphocytes T effecteurs contre diverses protéines musculaires, favorise la sécrétion de facteurs anti-inflammatoires et supprime l'inflammation chronique.
Effet : Ralentit la dégradation des protéines musculaires.
4.2 Mécanisme et effets de la promotion de la synthèse
Mécanisme : La poudre ultrafine de protéines musculaires non dénaturées contient des structures de réticulation covalentes primaires et d’ordre supérieur intactes. Les enzymes digestives intestinales peuvent sélectionner et décomposer spécifiquement les structures de réticulation covalentes primaires (liaisons peptidiques) tout en préservant les réticulations covalentes d’ordre supérieur. Il en résulte de petites molécules telles que des peptides réticulés et des peptides conjugués, qui régulent ensuite l’activité catalytique des protéases de multiples façons – en tant que substrats, chaperons moléculaires ou molécules de signalisation – afin de stimuler la synthèse des protéines musculaires. Une dégradation plus poussée en acides aminés individuels permet une participation directe à la synthèse des protéines musculaires.
Effet : Favorise la synthèse musculaire.
Cette régulation à double action — inhibant la dégradation tout en favorisant la synthèse — offre une efficacité nettement supérieure à celle des protéines ou des peptides dénaturés.
V. Dérivés de tissus
5.1 Structure déterminée à partir de tissus
Les principales fonctions de la nutrition non dénaturée (immunomodulation, régulation de l'activité) sont assurées par des structures naturellement actives. Ces structures présentent une spécificité élevée, n'exerçant d'effets régulateurs que sur les protéines ayant des structures identiques.
5.2 Origine spécifique aux tissus
Les ingrédients nutritionnels non dénaturés proviennent de tendons de bœuf ou de blancs de poulet, deux tissus musculaires squelettiques. Les tendons de bœuf présentent une similarité structurale de 95 %, tandis que les blancs de poulet présentent une similarité structurale de 85 %.
VI. Populations appropriées
6.1 Athlètes : Personnes pratiquant l’entraînement de force et le développement musculaire.
6.2 Sarcopénie : Inhibe la dégradation musculaire et favorise la synthèse musculaire.
Protéines spléniques non dénaturées – Santé immunitaire
I. Aperçu
La poudre ultrafine de protéines de rate non dénaturées est un ingrédient alimentaire fonctionnel non dénaturé, produit à partir de rate bovine fraîche. Elle est fabriquée selon une technologie de traitement ultrafin à très basse température (brevet d'invention national) et une technologie de séparation à très basse température basée sur le principe de l'enceinte à vent (technologie exclusive), conformément aux normes en vigueur.
II. Rapports et fonctions des composants

2.1 Composition et fonctions des complexes protéiques de la réponse immunitaire centrale
2.1.1 Protéines liées à l'immunité (complexe fonctionnel central) : La fraction la plus distinctive et fonctionnelle parmi les protéines de la rate.
Composition : Anticorps (immunoglobulines IgG, IgM, etc.) et leurs fragments.
Fonction : Acteurs principaux de l'immunité humorale.
2.1.2 Protéines du complexe majeur d'histocompatibilité
Composition : Molécules MHC de classe I et de classe II, largement exprimées à la surface des cellules présentatrices d'antigènes.
Fonction : Capturer les fragments de protéines pathogènes (antigènes) et les « présenter » aux lymphocytes T.
2.1.3 Protéines du système du complément
Composition : Plusieurs composants du complément, notamment C3, C4 et des activateurs du complément.
Fonction : Former le « complexe d’attaque membranaire du complément » pour perforer directement les membranes des agents pathogènes ; agir comme « chimiokines » pour attirer les cellules immunitaires sur les sites d’infection ; marquer les agents pathogènes pour favoriser la phagocytose (activité complémentaire).
2.1.4 Cytokines et chimiokines
Composition : Interleukines, interférons, facteurs de nécrose tumorale, etc.
Fonction : Servir de « molécules de signalisation » entre les cellules immunitaires, dirigeant avec précision l’initiation, l’amplification, la suppression ou la terminaison des réponses immunitaires et coordonnant le comportement de différents types de cellules immunitaires.
2.1.5 Récepteurs de reconnaissance de formes
Composition : Récepteurs de type Toll, etc.
Fonction : Reconnaître les motifs moléculaires conservés des agents pathogènes afin de déclencher des réponses immunitaires innées rapides.
2.2 Protéines impliquées dans le traitement des globules rouges
2.2.1 Enzymes de dégradation de l'hémoglobine
Composition : Hème oxygénase, biliverdine réductase, etc.
Fonction : Décomposer l'hémoglobine des globules rouges vieillissants, libérant des ions de fer pour leur réutilisation et convertissant l'hème en bilirubine.
2.2.2 Protéines de stockage et de transport du fer
Composition : La ferritine est le constituant le plus important.
Fonction : Stocke et stabilise le fer de manière sûre et efficace, empêchant ainsi le fer libre de générer des radicaux libres nocifs, et libère le fer selon les besoins de l'organisme.
2.3 Protéines structurales et d'échafaudage
Composition : Protéines de la matrice extracellulaire (par exemple, collagène, fibronectine) et molécules d'adhésion cellulaire.
Fonction : Former la structure physique de la pulpe blanche (gaines lymphatiques périartérielles, follicules lymphoïdes) et de la pulpe rouge de la rate ; assurer le ciblage, la rétention et les interactions des lymphocytes entre les cellules immunitaires.
2.4 Métabolisme et enzymes antioxydantes
2.4.1 Enzymes antioxydantes
Composition : Superoxyde dismutase, catalase, glutathion peroxydase.
Fonction : Éliminer les espèces réactives de l'oxygène (ROS) pour protéger les tissus spléniques des dommages oxydatifs.
2.4.2 Enzymes du métabolisme des nucléotides
Composition : Purine nucléoside phosphorylase, etc.
Fonction : Participe au métabolisme des purines au sein des lymphocytes, essentiel au fonctionnement des cellules immunitaires.
2.5 Transduction du signal et protéines régulatrices
Composition : Facteurs de transcription, notamment NF-κB, les protéines de la famille STAT, etc.
Fonction : Recevoir des signaux tels que des cytokines, se transloquer dans le noyau pour réguler l'expression de gènes spécifiques et déterminer le devenir d'activation, de prolifération ou de différenciation des cellules immunitaires.
III. Mécanisme d'action et effets
La poudre ultrafine de protéines spléniques non dénaturées fournit des protéines spléniques intactes, préservant leur conformation naturelle. Son mécanisme d'action principal ne repose pas sur l'apport de nutriments, mais plutôt sur sa fonction de réservoir naturel complexe de molécules bioactives. Elle fournit à l'organisme, et plus particulièrement au système immunitaire, des signaux régulateurs et des modules fonctionnels, exerçant ainsi un effet d'« immunité nutritionnelle ».
3.1 Effets immunitaires intestinaux locaux
3.1.1 Effets immunitaires passifs
Les particules ultrafines de protéines spléniques non dénaturées contiennent une grande quantité d'immunoglobulines qui atteignent l'intestin intactes grâce à une protection structurale. Leurs fragments de liaison aux antigènes (régions Fab) conservent leur activité pendant un certain temps, exerçant une immunité passive locale. Ces fragments Fab peuvent : - neutraliser directement des virus, des bactéries et leurs toxines spécifiques, les rendant incapables d'infecter ou d'induire une toxicité ; - adhérer aux pathogènes pour bloquer l'invasion ; - former des complexes avec les pathogènes ou les antigènes, stimulant ainsi le péristaltisme intestinal et facilitant leur encapsulation par le mucus, ce qui accélère leur expulsion par les selles et réduit leur temps de rétention et leur impact sur l'intestin.
3.1.2 Maintien de la fonction de barrière intestinale
Réduction des dommages inflammatoires : En éliminant rapidement les agents pathogènes et les toxines, il réduit les attaques directes sur les cellules épithéliales intestinales et la réponse inflammatoire excessive qui en résulte, protégeant ainsi l’intégrité de la barrière intestinale.
Soutien à la réparation : Certaines études suggèrent que d’autres composants des préparations d’immunoglobulines (tels que les facteurs de croissance) ou les anticorps eux-mêmes peuvent indirectement favoriser la réparation de la muqueuse.
3.1.3 Modulation de l'environnement immunitaire local
Protection antigénique : Au sein de l’intestin, le plus grand organe immunitaire du corps, un excès d’antigènes exogènes (alimentaires et pathogènes) peut déclencher des réactions immunitaires anormales. Les immunoglobulines orales se lient à une partie de ces antigènes et les éliminent, réduisant ainsi la charge pesant sur le système immunitaire intestinal et le risque d’attaques inappropriées, contribuant de ce fait au maintien de l’équilibre de la tolérance immunitaire.
Effets anti-inflammatoires : En réduisant la stimulation des agents pathogènes par les mécanismes susmentionnés, il diminue la production de cytokines pro-inflammatoires, créant un environnement local plus propice à la réparation des tissus et à l’équilibre microbien.
3.2 Effets immunomodulateurs systémiques
3.2.1 Induction de la tolérance immunitaire périphérique par les antigènes
La rate contient des protéines et des antigènes liés à l'immunité sous leur forme native, non dénaturée. Ces substances peuvent être reconnues par le système immunitaire intestinal comme des « stimuli immunitaires familiers et bénéfiques », renforçant ainsi la vigilance et la réactivité immunitaires sans déclencher d'inflammation intense (voie de tolérance orale).
3.2.2 Fourniture directe de composants immunologiquement actifs et de molécules de signalisation
Libère des peptides à petites molécules dotés d'une activité immunomodulatrice au sein de l'intestin. Ces fragments peptidiques agissent comme des molécules de signalisation reconnues par les cellules immunitaires intestinales (par exemple, celles des plaques de Peyer), stimulant et entraînant en douceur le système immunitaire, à la manière d'une « répétition immunitaire ».
3.2.3 Interaction avec le référentiel d'informations immunitaires
Potentiel d'induction de la tolérance orale : pour certaines prédispositions auto-immunes, des théories suggèrent que l'ingestion de tissus contenant des antigènes similaires à ceux de l'organisme pourrait contribuer à induire une tolérance immunitaire.
IV. Dérivés de tissus
4.1 Dépendance à la structure dérivée des tissus
Les principales fonctions de la nutrition non dénaturée (immunomodulation, régulation de l'activité) sont assurées par des structures naturellement actives. Ces structures présentent une spécificité élevée, n'exerçant d'effets régulateurs que sur les protéines ayant des structures identiques.
4.2 Origine du tissu correspondant
La rate, riche en cellules et facteurs immunitaires divers, correspond au tissu immunitaire de par son origine tissulaire.
V. Populations appropriées
5.1 Personnes immunodéprimées
5.2 Personnes atteintes de troubles digestifs
Élastine non dénaturée – Santé vasculaire
I. Aperçu
La poudre ultrafine d'élastine non dénaturée est un ingrédient alimentaire fonctionnel non dénaturé, produit à partir de ligaments nucaux ou d'aortes bovines fraîches. Sa fabrication est conforme aux normes en vigueur et utilise une ultramicronisation à très basse température (technologie brevetée) et une technologie de séparation à très basse température basée sur le principe de l'enceinte à vent (technologie exclusive).
II. Proportions des composants
2.1 Composition de la poudre ultrafine d'élastine non dénaturée (ligament nuchal bovin) et de sa poudre lyophilisée

2.2 Composition de la poudre ultrafine d'élastine non dénaturée (aorte bovine) et de sa poudre lyophilisée

III. Structure active naturelle
3.1 Structure primaire
La séquence d'acides aminés de l'élastine est constituée de segments hydrophobes alternés (riches en valine, alanine, glycine et proline) et de segments réticulés (riches en lysine et alanine) disposés en tandem.
3.2 Structure supérieure
La séquence désordonnée d'acides aminés de la protéine élastine empêche la formation de structures secondaires régulières, dépourvues de structures conventionnelles de niveaux II, III ou IV. Elle forme en revanche un réseau polymère tridimensionnel désordonné par le biais de liaisons covalentes, telles que les pontages élastiques.
IV. Mécanisme d'action et effets
L'élastine est une protéine non nutritionnelle dont la fonction principale n'est pas l'apport d'acides aminés, mais plutôt sa « structure active naturelle » unique, qui exerce des effets structuraux.
4.1 Mécanisme et effets de la régulation immunitaire périphérique
4.1.1 Mécanisme : La structure multidimensionnelle unique de l’élastine non dénaturée active la différenciation des lymphocytes T des plaques de Peyer en lymphocytes T régulateurs spécifiques (Tregs). Ceci module l’attaque erronée du système immunitaire contre l’élastine endogène et supprime les réponses inflammatoires.
4.1.2 Effets : Ralentit la dégradation de l’élastine. Avec l’âge (à partir de 25-30 ans environ), la capacité de l’organisme à synthétiser l’élastine diminue fortement. Parallèlement, en raison d’un dérèglement immunitaire, les fibres d’élastine existantes se fracturent, se rigidifient et deviennent dysfonctionnelles par glycation, stress oxydatif (dommages causés par les radicaux libres) et dégradation enzymatique (par exemple, par les métalloprotéinases matricielles). Il s’agit de la principale cause de l’athérosclérose chez l’être humain.
4.2 En tant que molécule de signalisation cellulaire (fonction la plus notable)
4.2.1 Mécanisme : La surface des molécules d’élastine intactes contient des domaines bioactifs spécifiques (par exemple, des domaines de liaison cellulaire, des domaines chimiotactiques). Lorsque ces domaines conservent leur conformation native, ils peuvent se lier spécifiquement à des récepteurs (par exemple, des protéines de liaison à l’élastine) présents à la surface des fibroblastes, des cellules endothéliales et d’autres cellules. Cette liaison stimule la synthèse de collagène, d’élastine et d’acide hyaluronique supplémentaires par les cellules. Ce processus induit une régénération endogène, bien plus importante qu’un simple apport de matières premières.
4.2.2 Effet : Contribue à améliorer fondamentalement l'élasticité et la fermeté, restaurant l'élasticité des tissus tels que les vaisseaux sanguins.
4.2 En tant que substrat modulant l'activité enzymatique
4.2.1 Mécanisme : L’élastine non dénaturée peut être dégradée dans l’intestin en peptides réticulés contenant des structures de réticulation covalentes primaires et secondaires. Ces peptides réticulés servent de substrats aux enzymes de modification structurale concernées, ce qui augmente leur activité catalytique.
4.2.2 Effet : Favorise la formation ou la réparation des structures d'élastine.
4.3 En tant que gabarit structurel et échafaudage
4.3.1 Mécanisme : L’élastine non dénaturée, conservant de grands fragments structuraux et des structures réticulées non dénaturées, peut servir de « structure de guidage » in vitro ou in vivo. Elle oriente les molécules précurseurs d’élastine nouvellement synthétisées vers un agencement ordonné et une réticulation correcte, facilitant ainsi la formation de nouveaux réseaux de fibres élastiques fonctionnels.
4.3.2 Effet : Aide à réparer les fibres élastiques cassées et désordonnées causées par le photovieillissement ou le vieillissement.
V. Dérivés de tissus
5.1 Dépendance à la structure dérivée des tissus
Les fonctions des protéines non dénaturées (immunomodulation, régulation de l'activité) dépendent de leur structure. Celle-ci présente une spécificité élevée, ne régulant que les protéines de structure identique.
5.2 Sources tissulaires correspondantes
Les tissus riches en élastine, tels que les vaisseaux sanguins, la peau et les alvéoles.
VI. Candidats appropriés
6.1 Santé vasculaire : Hypertension, hyperlipidémie, athérosclérose.
6.2 Soin anti-rides pour la peau : Relâchement cutané, rides, perte d'élasticité.
6.3 Insuffisance pulmonaire : Patients atteints d’emphysème, de bronchopneumopathie chronique obstructive (BPCO), etc.





