Неденатурированные костные ткани: питание и здоровье костей.
I. Обзор
В нашу линейку неденатурированных питательных продуктов для здоровья костей входят: порошок для питания целых костей , ультрамикропорошок для питания костей , ультрамикропорошок кальция с неденатурированным костным белком и неденатурированный ультрамикропорошок коллагена I типа . Эти продукты изготавливаются из свежей, соответствующей возрасту коровьей бедренной кости (включая суставы) в качестве основного сырья, что обеспечивает высочайшее качество коллагена, гликозаминогликанов и костных белков. Производственный процесс включает в себя сверхнизкотемпературную ультрамикроизацию и технологию сверхнизкотемпературного разделения на основе принципа воздушного потока , что гарантирует сохранение неденатурированной структуры . Это сохраняет функциональные свойства ингредиентов, делая их идеальными для поддержания здоровья костей и восстановления тканей.
Уникальная неденатурированная природа этих продуктов гарантирует сохранение их естественной активной структуры, что крайне важно для полного усвоения организмом биоактивных соединений. Эта инновационная технология обеспечивает превосходное усвоение и эффективность в поддержании здоровья костей и суставов.
II. Соотношения компонентов

*Коллаген в основном состоит из коллагена I типа (более 90%), с небольшим количеством коллагена II типа (менее 10%).
III. Естественная активная структура
3.1 Первичная структура
3.1.1 Коллаген I типа: преимущественно состоит из повторяющихся аминокислотных последовательностей Gly-Pro-X или Gly-X-Hyp (пептидных связей).
3.1.2 Гликозаминогликаны
Гиалуроновая кислота (ГК): простая повторяющаяся линейная полисахаридная цепь, образованная чередующимися гликозидными связями β(1→3) и β(1→4) между D-глюкуроновой кислотой и N-ацетилглюкозамином.
Хондроитинсульфат (ХС): серия микроскопически гетерогенных линейных полисахаридных цепей с основной структурой из чередующихся D-глюкуроновой кислоты и N-ацетилгалактозамина, характеризующихся сложными и вариабельными модификациями сульфатирования в определенных положениях на N-ацетилгалактозамине (и иногда на глюкуроновой кислоте).
3.2 Высшие структуры
3.2.1 Коллаген I типа
Вторичная структура: α-пептидная цепь имеет левостороннюю спиральную структуру (в основном водородные связи).
Третичная структура: Тройная спираль (пептидные связи, водородные связи, силы Ван дер Ваальса, гидрофобные взаимодействия, ковалентные сшивки, образующиеся в результате альдольной конденсации).
3.2.2 Высшие структуры гликозаминогликанов (ГАГ)
Высшая структура гиалуроновой кислоты: в растворе образует хаотично свернутую, сильно гидратированную сеть.
Высшая структура хондроитинсульфата: ковалентно связан с основными белками, образуя протеогликаны.
3.3 Надструктура (супрамолекулярная ковалентная сшивка)
Между макромолекулами коллагена I типа: структура пиридинолиновых поперечных связей.
Между остеокальцином и гидроксиапатитом (ГАП): кальций, хелатированный γ-карбоксиглутаминовой кислотой.
IV. Механизм действия и эффекты
Порошок для питания целых костей, ультрадисперсный неденатурированный порошок для питания костей, ультрадисперсный неденатурированный костный белок и кальций, а также ультрадисперсный неденатурированный коллаген I типа не только обеспечивают комплексное, сбалансированное питание, необходимое для роста и восстановления костной ткани, но и приносят пользу благодаря наличию «активных структур».
4.1 Механизм и эффекты усиления всасывания кальция
4.1.1 Механизм: Сверхковалентная структура (кальций, хелатированный γ-карбоксиглутаминовой кислотой) между остеокальцином в неденатурированном костном белке, ультрадисперсном порошке кальция и гидроксиапатитом (ГАП) защищена трехмерной структурой белка, предотвращая контакт с желудочной кислотой. Это позволяет комплексу достигать кишечника в неизмененном виде. Под действием кишечных пищеварительных ферментов он расщепляется на растворимые молекулы кальция, такие как кальций, хелатированный γ-карбоксиглутаминовой кислотой, или кальций, связанный с низкомолекулярными пептидами. Это увеличивает концентрацию молекул кальция в кишечнике, обеспечивая всасывание по парацеллюлярному пути, обусловленному градиентами концентрации.
4.1.2 Эффект: Эта абсорбция основана на градиентах концентрации и не зависит от насыщения кальцийсвязывающим белком (CaBP). У пациентов со сниженным уровнем CaBP вследствие роста, развития или старения, что приводит к ограниченному усвоению кальция, это обеспечивает физиологическую компенсацию поглощения кальция.
4.2 Механизм и эффект стимуляции остеогенеза с помощью кальция
4.2.1 Механизм: Остеогенез кальция регулируется множеством факторов как в крови, так и в костной ткани.
Кровяная среда в первую очередь зависит от коэффициента произведения концентрации кальция и фосфора. Неденатурированный ультрадисперсный порошок кальция из костного белка постепенно переваривается и расщепляется в кишечнике на растворимые молекулы кальция, такие как кальций, хелатированный γ-карбоксиглутаминовой кислотой, или кальций, связанный с низкомолекулярными пептидами, что обеспечивает медленное всасывание. Соотношение кальция и фосфора приближается к 2:1, что максимизирует произведение концентрации кальция и фосфора и способствует притоку кальция из крови в костную ткань.
На состояние костной ткани влияют три фактора: нанообласть гидроксиапатита (HAP) коллагена I типа, карбоксилирование остатков глутаминовой кислоты в остеокальцине, а также регуляция со стороны связующих белков и молекул адгезии костной ткани.
Тройная спиральная структура содержащихся в нем неденатурированных фрагментов коллагена I типа может активировать кишечную лимфатическую систему, регулируя иммунитет, подавляя воспалительные реакции и замедляя деградацию коллагена I типа. Продукты его деградации — небольшие молекулы, такие как сшитые пептиды и конъюгированные пептиды, — индуцируют образование структуры коллагена I типа посредством множественных механизмов (субстрат, молекулярный шаперон, сигнализация). Далее, расщепляясь на мономеры аминокислот, они непосредственно участвуют в синтезе коллагена, обеспечивая достаточное количество нанообластей для ГАП и максимизируя нанообласти коллаген-ГАП.
Его неденатурированные костные белки связывают кальций, образуя кальций-связанную γ-карбоксиглутаминовую кислоту или кальций-связанные небольшие пептиды. В качестве альтернативы, они активируют γ-карбоксиглутамилкарбоксилазу в качестве субстратов или посредством передачи сигнала, катализируя карбоксилирование трех остатков глутаминовой кислоты в остеокальцине. Это обеспечивает места связывания кальция, максимизируя точки прикрепления кальция.
Содержащиеся в нем неденатурированный десмин и неденатурированный остеонектин могут способствовать синтезу соответствующих белков, пространственно ограничивать присутствие ГАП внутри коллагена I типа, закреплять коллаген на ГАП и предотвращать потерю кальция в костях.
4.2.2. Эффект: Кальций может проникать в клетки, накапливаться и оставаться в них.
4.3 Механизм и эффекты повышения содержания белка в костной ткани
К костным белкам относятся коллаген (в основном коллаген I типа) и гликопротеины (главным образом остеокальцин, фибронектин, остеонектин и более 10 других). Механизм увеличения их содержания принципиально схож, что иллюстрируется примером неденатурированного ультрадисперсного порошка коллагена I типа.
4.3.1 Механизм
Механизм регуляции кишечного иммунитета: ультрадисперсный порошок неденатурированного коллагена I типа обладает трехмерной структурой, устойчивой к разрушению желудочным соком, что обеспечивает прямой физический контакт с лимфатической системой кишечника. Это активирует незрелые Т-клетки, которые дифференцируются в специфические регуляторные Т-клетки (Treg), подавляя ошибочную атаку иммунной системы на коллаген I типа. Это способствует секреции противовоспалительных факторов, подавляя хроническое воспаление и замедляя деградацию коллагена I типа.
Механизм регуляции активности внутриклеточных ферментов: ультрадисперсный порошок неденатурированного коллагена I типа содержит неповрежденные первичные, высоко- и сверхковалентные сшивки. Кишечные пищеварительные ферменты избирательно расщепляют первичные (пептидные) ковалентные сшивки, сохраняя при этом высоко- и сверхковалентные сшивки. Этот процесс высвобождает небольшие молекулы, такие как сшитые пептиды и конъюгированные пептиды, или служит субстратами, молекулярными шаперонами или сигнальными молекулами для индукции структурного формирования коллагена I типа.
Обеспечение питательными веществами: Сшитые пептиды, конъюгированные пептиды и другие малые молекулы далее расщепляются на отдельные аминокислоты, непосредственно участвуя в синтезе коллагена.
4.3.2 Эффекты
Такое двойное действие — ингибирование деградации и одновременное стимулирование синтеза — обеспечивает значительно большую эффективность, чем роль денатурированных белков или пептидов в обеспечении питательными веществами.
4.4 Механизм и эффекты усиления действия гликозаминогликанов (ГАГ)
4.4.1 Механизм
Неденатурированные ультрадисперсные частицы гликозаминогликанов доставляют фрагменты ГК, ХС и КС. Они легко расщепляются на низкомолекулярные фрагменты, олигосахариды и дисахаридные единицы. Они активируют хондроциты, ингибируют ферменты, расщепляющие матрикс (ММП или АДАМТС), регулируют иммунитет и оказывают противовоспалительное и анальгетическое действие. Внутриклеточно они далее расщепляются на отдельные молекулы гликозаминогликанов, непосредственно участвуя в синтезе ГК, ХС и КС.
4.4.2 Эффекты
Повышает содержание гликозаминогликанов.
V. Тканеобразные материалы
5.1. Происхождение из ткани определяется структурой.
Основные функции неденатурированного питания (иммуномодуляция, регуляция активности ферментов) достигаются благодаря его естественной активной структуре. Эта структура обладает высокой специфичностью, определяя свойства, присущие тканям.
5.2 Тканевая природа неденатурированных питательных веществ для костей
Порошок для питания целых костей, ультрадисперсный порошок для питания неденатурированных костей, ультрадисперсный порошок костного белка кальция и ультрадисперсный порошок коллагена I типа используют в качестве сырья свежие, соответствующие возрасту кости голени крупного рогатого скота (включая суставы). Их основным белком является коллаген I типа, и их происхождение из ткани следует определять на основе коллагена I типа. Хотя коллаген I типа в кожной ткани имеет генетическую гомологию с костной тканью и идентичные аминокислотные последовательности, различия в микроокружающих модификациях приводят к различным высокоуровневым и ультраструктурным структурам. Поэтому кожная ткань не считается гомологичной серии неденатурированных порошков для питания костей. К основным гомологичным тканям относятся: твердая костная ткань, межпозвоночные диски, мениски и небольшое количество суставного хряща.
VI. Подходящая популяция
6.1 Потребности в питательных веществах: Подходит для всех людей с дефицитом кальция, особенно для младенцев, маленьких детей и пожилых людей.
6.2 Потребности в росте костей: Подходит для тех, кому необходимо увеличить рост на этапах развития, и для тех, кто стремится замедлить потерю роста в процессе старения.
6.2 Потребности в увеличении костной массы
6.2.1. Лица с симптомами рахита в период роста и развития: такие как задержка закрытия родничка, квадратный родничок, плоская голова, голубиная грудная клетка, воронкообразная грудная клетка, бугорчатые изменения, вальгусная или варусная деформация коленных суставов, сколиоз и т. д.
6.2.2 Лица среднего и пожилого возраста: лица, у которых наблюдаются признаки гиперплазии костной ткани, уменьшение роста, кифоз, остеопороз, грыжи межпозвоночных дисков и т. д.
6.3 Потребности в восстановлении при дегенеративных изменениях суставов: Пациенты с дегенеративным остеоартрозом, особенно те, у кого имеются повреждения или воспаление мениска.
6.4 Потребности в заживлении костных травм: Заживление костных травм или рост костного цемента после имплантации и т. д.
Питание при недегенеративных заболеваниях суставов — Серия статей о здоровье суставов
I. Обзор
В серию неденатурированных пищевых добавок для здоровья суставов входят: порошок для полного питания суставов, ультрадисперсный порошок для питания суставов без денатурации, ультрадисперсный порошок для питания хрящей без денатурации и ультрадисперсный порошок коллагена II типа без денатурации. Все они являются функциональными пищевыми ингредиентами, произведенными из свежих, соответствующих возрасту бычьих коленных суставов или хрящей. Производство осуществляется в соответствии с соответствующими стандартами с использованием сверхнизкотемпературной сверхтонкой обработки (запатентованная технология национального изобретения) и технологии сверхнизкотемпературного разделения по принципу ветрокамеры (собственная технология).
II. Соотношения компонентов

*Коллаген в основном состоит из коллагена II типа (более 90%), с небольшим количеством коллагена I типа (менее 10%).
III. Естественная активная структура
3.1 Первичная структура
3.1.1 Коллаген II типа: преимущественно состоит из повторяющихся аминокислотных последовательностей Gly-Pro-X или Gly-X-Hyp (пептидная связь).
3.1.2 Гликозаминогликаны
Гиалуроновая кислота (ГК): простая повторяющаяся линейная полисахаридная цепь, образованная чередующимися гликозидными связями β(1→3) и β(1→4) между D-глюкуроновой кислотой и N-ацетилглюкозамином.
Хондроитинсульфат (ХС): серия микроскопически гетерогенных линейных полисахаридных цепей, образованных чередующимися звеньями D-глюкуроновой кислоты и N-ацетилгалактозамина в качестве основной цепи, со сложными и вариабельными модификациями сульфатирования, происходящими в определенных положениях на звеньях N-ацетилгалактозамина (и иногда глюкуроновой кислоты).
3.2 Высшие структуры
3.2.1 Коллаген II типа
Вторичная структура: α-пептидная цепь имеет левостороннюю спиральную структуру (в основном водородные связи).
Третичная структура: Тройная спираль (пептидные связи, водородные связи, силы Ван дер Ваальса, гидрофобные взаимодействия, ковалентные сшивки, образующиеся в результате альдольной конденсации).
3.2.2 Гликозаминогликаны (ГАГ): структура высшего порядка
Высшая структура гиалуроновой кислоты: в растворе образует хаотично свернутую, сильно гидратированную сетевую структуру.
Высокоуровневая структура хондроитинсульфата: ковалентно связана с основными белками, образуя протеогликаны.
3.3 Надмолекулярная структура (надмолекулярная ковалентная сшивка)
Межмолекулярное сшивание коллагена II типа: гидроксипиридолин.
Остеокальцин и гидроксиапатит (ГАП): γ-карбоксиглутаминовая кислота образует хелатные комплексы с кальцием.
IV. Механизм действия и эффекты
Комплексный питательный порошок для суставов, неденатурированный ультрадисперсный питательный порошок для суставов, неденатурированный ультрадисперсный питательный порошок для хрящевой ткани и неденатурированный ультрадисперсный порошок коллагена II типа не только обеспечивают комплексное, сбалансированное питание для суставных тканей, но и оказывают «активное структурное» воздействие.
4.1 Механизм и эффекты стимуляции усвоения кальция
4.1.1 Механизм: Сверхковалентная структура (кальций, хелатированный γ-карбоксиглутаминовой кислотой) между остеокальцином и гидроксиапатитом (ГАП) в неденатурированном ультрадисперсном порошке костного белка кальция предотвращает контакт с желудочным соком благодаря защите трехмерной структуры белка. Это позволяет ему достигать кишечника в неизмененном виде, где он расщепляется кишечными пищеварительными ферментами на растворимые молекулы кальция, такие как кальций, хелатированный γ-карбоксиглутаминовой кислотой, или кальций, связанный с низкомолекулярными пептидами, тем самым увеличивая концентрацию кальция в кишечнике. Под действием градиента концентрации кальций абсорбируется парацеллюлярным путем.
4.1.2 Эффект: Эта абсорбция основана на градиентах концентрации и не зависит от насыщения кальцийсвязывающим белком (CaBP). У пациентов со сниженным уровнем CaBP вследствие роста, развития или старения, что приводит к ограниченному усвоению кальция, это обеспечивает физиологическую компенсацию поглощения кальция.
4.2 Механизм и эффект стимуляции остеогенеза с помощью кальция
4.2.1 Механизм: Остеогенез кальция регулируется множеством факторов как в крови, так и в костной ткани.
Кровяная среда в первую очередь зависит от коэффициента произведения концентрации кальция и фосфора. Неденатурированный ультрадисперсный порошок кальция из костного белка постепенно переваривается и расщепляется в кишечнике на растворимые молекулы кальция, такие как кальций, хелатированный γ-карбоксиглутаминовой кислотой, или кальций, связанный с низкомолекулярными пептидами, что обеспечивает медленное всасывание. Соотношение кальция и фосфора приближается к 2:1, что максимизирует произведение концентрации кальция и фосфора и способствует притоку кальция из крови в костную ткань.
На состояние костной ткани влияют три фактора: нанообласть HAP коллагена I типа, карбоксилирование остатков глутаминовой кислоты в остеокальцине, а также регуляция с помощью бриджина и остеонектина.
Неденатурированные фрагменты коллагена I типа, содержащиеся в нем, активируют кишечную лимфатическую систему, модулируя иммунитет, подавляя воспалительные реакции и замедляя деградацию коллагена I типа. Продукты его деградации — небольшие молекулы, такие как сшитые пептиды и конъюгированные пептиды, — функционируют как субстраты, молекулярные шапероны и сигнальные пути, индуцирующие структурное формирование коллагена I типа. Дальнейшая деградация до аминокислотных мономеров непосредственно участвует в синтезе коллагена, обеспечивая достаточное количество нанообластей для гидроксиапатита и максимизируя нанообласти коллагена.
Его неденатурированные костные белки связывают кальций, образуя кальций-связанную γ-карбоксиглутаминовую кислоту или кальций-конъюгированные небольшие пептиды. В качестве альтернативы, они активируют γ-карбоксиглутаминовую кислоту карбоксилазу в качестве субстратов или посредством передачи сигнала, катализируя карбоксилирование трех остатков глутаминовой кислоты в остеокальцине. Это обеспечивает места связывания кальция, максимизируя точки прикрепления кальция.
Содержащиеся в нем неденатурированный десмин и неденатурированный остеонектин могут способствовать синтезу соответствующих белков, пространственно ограничивать присутствие ГАП внутри коллагена I типа, закреплять коллаген на ГАП и предотвращать потерю кальция в костях.
4.2.2. Эффект: Кальций поступает, задерживается и остается в организме.
4.3 Механизм и эффекты восстановления хрящевой ткани
Суставной хрящ представляет собой гиалиновый хрящ, основным белком которого является коллаген II типа.
4.3.1 Механизм
Механизм регуляции кишечного иммунитета: Неденатурированный ультрадисперсный порошок коллагена II типа обладает трехмерной структурой, которая препятствует деградации его естественной активной формы под воздействием желудочного сока. Он достигает кишечника и физически взаимодействует со специфической кишечной лимфатической системой, активируя наивные Т-клетки, которые трансформируются в специфические регуляторные Т-клетки (Treg). Treg-клетки модулируют эффекторные Т-клетки, предотвращая ошибочные иммунные атаки на коллаген II типа, секретируют противовоспалительные факторы для подавления хронического воспаления и замедляют деградацию коллагена II типа.
Механизм регуляции активности ферментов внутри органов: ультрадисперсный порошок неденатурированного коллагена II типа содержит неповрежденные первичные, высокоуровневые и суперуровневые ковалентные сшивающие структуры. Кишечные пищеварительные ферменты избирательно расщепляют первичные (пептидные) ковалентные сшивки, сохраняя при этом высокоуровневые и суперуровневые структуры. В результате образуются малые молекулы, такие как сшитые пептиды и конъюгированные пептиды, или молекулы, которые могут служить субстратами, молекулярными шаперонами или сигнальными молекулами, индуцирующими формирование структуры коллагена II типа.
Действие питательных веществ: Небольшие молекулы, такие как сшитые пептиды и конъюгированные пептиды, далее расщепляются на аминокислоты, которые участвуют в синтезе коллагена II типа.
4.3.2 Эффекты
Такое двойное действие — ингибирование деградации и одновременное стимулирование синтеза — обеспечивает значительно большую эффективность, чем эффекты, опосредованные денатурированными белками или пептидами.
4.4 Механизм действия и эффекты гликозаминогликанов (ГАГ)
4.4.1 Механизм
Неденатурированные ультрадисперсные частицы ГАГ содержат фрагменты ГК, хондроитинсульфата и хондроитинсульфата, которые легко расщепляются на низкомолекулярные фрагменты, олигосахариды и дисахаридные единицы. Эти фрагменты активируют хондроциты, ингибируют ферменты, расщепляющие матрикс (ММП или АДАМТС), регулируют иммунитет и оказывают противовоспалительное и обезболивающее действие. Кроме того, они могут дополнительно расщепляться внутриклеточно на отдельные молекулы гликозаминогликанов, непосредственно участвуя в синтезе ГК, хондроитинсульфата и хондроитинсульфата.
4.4.2 Эффекты
Повышает содержание гликозаминогликанов.
V. Тканевые материалы
5.1. Происхождение из ткани определяется структурой.
Основные функции неденатурированного питания (иммуномодуляция, регуляция активности ферментов) достигаются благодаря его естественной активной структуре. Эта структура обладает высокой специфичностью, определяя свойства, присущие тканям.
5.2 Тканевая природа неденатурированных питательных смесей для суставов
В состав комплексных питательных смесей для суставов, ультрадисперсных неденатурированных питательных смесей для суставов, ультрадисперсных неденатурированных питательных смесей для хрящевой ткани и ультрадисперсных неденатурированных коллагенов II типа входят свежие, соответствующие возрасту бычьи коленные суставы или хрящи. Их основным белком является коллаген II типа, тканевое происхождение которого связано с хрящевой тканью, богатой коллагеном II типа. К основным гомологичным тканям относятся: суставной хрящ и небольшое количество твердой кости.
VI. Подходящая популяция
6.1 Потребности в восстановлении после травм суставов
6.1.1 Потребности в лечении дегенеративного остеоартроза
6.1.2 Потребности в восстановлении суставов после травм
Неденатурированное питание для кожи — серия статей о здоровье кожи.
I. Обзор
В серию неденатурированных питательных продуктов для здоровья кожи входят: порошок для питания цельной кожи, ультрадисперсный порошок для питания кожи без денатурации и ультрадисперсный порошок коллагена III типа без денатурации. Все они производятся из свежих, соответствующих возрасту шкур крупного рогатого скота или ослов. Изготовленные в соответствии с действующими стандартами с использованием ультрамикронизации при сверхнизких температурах (запатентованная технология национального изобретения) и технологии сверхнизкотемпературного разделения по принципу ветрокамеры (собственная технология), эти неденатурированные функциональные пищевые ингредиенты являются неденатурированными.
II. Соотношение ингредиентов

III. Природные активные структуры
3.1 Первичная структура
3.1.1 Коллаген типа I/III: преимущественно повторяющиеся аминокислотные последовательности Gly-Pro-X или Gly-X-Hyp (пептидные связи)
3.1.2 Гликозаминогликаны
Гиалуроновая кислота (ГК): простая повторяющаяся линейная полисахаридная цепь, образованная чередующимися гликозидными связями β(1→3) и β(1→4) между D-глюкуроновой кислотой и N-ацетилглюкозамином.
Хондроитинсульфат (ХС): серия микроскопически гетерогенных линейных полисахаридных цепей, образованных чередующимися звеньями D-глюкуроновой кислоты и N-ацетилгалактозамина в качестве основной цепи, со сложными и вариабельными модификациями сульфатирования, происходящими в определенных положениях на звеньях N-ацетилгалактозамина (и иногда глюкуроновой кислоты).
3.2 Высшие структуры
3.2.1 Коллаген типа I/III
Вторичная структура: α-пептидная цепь имеет левостороннюю спиральную структуру (в основном водородные связи).
Третичная структура: Тройная спираль (пептидные связи, водородные связи, силы Ван дер Ваальса, гидрофобные взаимодействия, ковалентные сшивки, образующиеся в результате альдольной конденсации).
3.2.2 Высшие структуры гликозаминогликанов (ГАГ)
Высшая структура гиалуроновой кислоты: в растворе образует хаотично свернутую, сильно гидратированную сеть.
Высшая структура хондроитинсульфата: основные белки ковалентно связываются, образуя протеогликаны.
3.3 Надструктура (супрамолекулярная ковалентная сшивка)
Межмолекулярное сшивание коллагена I типа: гидроксилизинпиридинолин и лизинпиридинолин.
IV. Механизм действия и эффекты
Порошок для питания всей кожи, ультрадисперсный неденатурированный порошок для питания кожи и ультрадисперсный неденатурированный коллаген III типа не только обеспечивают комплексное, сбалансированное питание, необходимое для роста и восстановления кожных тканей, но и оказывают воздействие «активных структур».
4.1 Механизм и эффекты усиления выработки коллагена
4.1.1 Механизм
Механизм регуляции кишечного иммунитета: ультрадисперсный порошок неденатурированного коллагена I/III типа обладает трехмерной структурой, которая препятствует деградации его естественной активной структуры под воздействием желудочного сока. Он достигает кишечника и физически взаимодействует с кишечной лимфатической системой, активируя наивные Т-клетки, которые трансформируются в специфические регуляторные Т-клетки (Treg). Treg модулируют эффекторные Т-клетки, предотвращая ошибочные иммунные атаки против неденатурированного коллагена I/III типа, секретируют противовоспалительные факторы для подавления хронического воспаления и замедляют деградацию неденатурированного коллагена I/III типа.
Механизм регуляции активности внутриклеточных ферментов: Неденатурированный ультрадисперсный порошок коллагена I/III содержит неповрежденные первичные, высоко- и суперковалентные сшитые структуры. Кишечные пищеварительные ферменты могут специфически отбирать и расщеплять первичные (пептидные) ковалентные сшитые структуры, сохраняя при этом высоко- и суперковалентные сшитые структуры. В результате этого процесса образуются малые молекулы, такие как сшитые пептиды и конъюгированные пептиды, которые затем индуцируют образование структур коллагена типа I/III посредством множественных путей, включая выполнение функций субстратов, молекулярных шаперонов или сигнальных молекул.
Функция питательных веществ: Небольшие молекулы, такие как сшитые пептиды и конъюгированные пептиды, могут далее расщепляться на отдельные молекулы аминокислот, непосредственно участвуя в синтезе коллагена.
4.1.2 Эффект
Такое двойное действие — ингибирование деградации и одновременное стимулирование синтеза — обеспечивает значительно большую эффективность, чем питательная функция, выполняемая денатурированными белками или пептидами.
4.2 Механизм и эффект усиления действия гликозаминогликанов (ГАГ)
4.2.1 Механизм
Неденатурированные ультрадисперсные частицы гликозаминогликанов содержат фрагменты гиалуроновой кислоты (ГК) и хондроитинсульфата (ХС), которые легко расщепляются на низкомолекулярные фрагменты, олигосахариды и дисахаридные единицы. Эти фрагменты ингибируют матрикс-деградирующие ферменты (ММП или АДАМТС), регулируют иммунные реакции, оказывая противовоспалительное и анальгетическое действие, а также могут дополнительно расщепляться внутриклеточно на отдельные молекулы гликозаминогликанов, непосредственно участвуя в синтезе ГК и ХС.
4.2.2 Эффекты
Повышает содержание гликозаминогликанов.
V. Свойства, полученные из тканей
5.1 Свойства тканей, определяемые структурой
Основные функции неденатурированного питания (иммуномодуляция, регуляция активности ферментов) достигаются благодаря его естественной активной структуре. Эта структура обладает высокой специфичностью, а её состав определяет свойства, присущие тканям.
5.2 Свойства неденатурированных питательных веществ для кожи, получаемые из тканей
Порошок для питания цельной кожи, ультрадисперсный неденатурированный порошок для питания кожи и ультрадисперсный неденатурированный коллаген III типа — все они используют в качестве сырья свежую, соответствующую возрасту ослиную или коровью кожу. Их основными белками являются коллаген I и III типов, а происхождение из ткани — кожная ткань, богатая этими типами коллагена. Основная гомологичная ткань: кожная ткань.
VI. Подходящие популяции
6.1 Антивозрастные средства для кожи
6.2 Восстановление повреждений кожи
Неденатурированное питание мозга - здоровье мозга
I. Обзор
Неденатурированный нейротрофический ультрадисперсный порошок из головного мозга — это неденатурированный функциональный пищевой ингредиент, производимый из свежего говяжьего или свиного мозга. Он изготавливается в соответствии с соответствующими стандартами с использованием сверхнизкотемпературного сверхтонкого измельчения (запатентованная технология национального изобретения) и технологии сверхнизкотемпературного разделения по принципу воздушной камеры (собственная технология).
II. Соотношения компонентов
2.1 Общая классификация
Вода, липиды, белки, неорганические соли, другие ключевые молекулы. Неорганические соли и ключевые молекулы присутствуют в очень низких концентрациях.

2.2 Белковый состав
Белки головного мозга составляют приблизительно 30% от сухого веса и могут быть классифицированы по функциям:
2.2.1 Структурные белки: белки микротрубочек, микрофиламенты (актин), нейрофиламентные белки.
2.2.2 Белки, ассоциированные с синапсами: белки ионных каналов, белки рецепторов нейромедиаторов, белки, ассоциированные с везикулами, белки-скелеты.
2.2.3 Белки, ассоциированные с миелином: основной белок миелина, протеолипидные белки, гликопротеины, ассоциированные с миелином.
2.2.4 Белки, участвующие в метаболизме и энергетическом обмене: митохондриальные белки, гексокиназа, энолаза
2.2.5 Нейротрофические факторы:
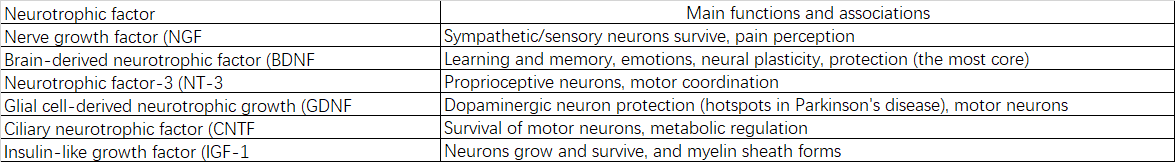
2.2.6 Сигнальные белки: G-белки, протеинкиназы, фосфатазы и др.
2.2.7 Белки, связанные с заболеванием: β-амилоид, тау-белок, α-синуклеин.
2.3 Липидный состав
2.3.1 Фосфолипиды: цереброзид, фосфатидилинозитол, фосфатидилсерин, сфингомиелин, лецитин и др.
2.3.2 Холестерин
2.2.3 Нейтральные жиры
В составе фосфолипидов и нейтральных жиров присутствуют специфические жирные кислоты: насыщенные жирные кислоты составляют приблизительно 30% и более , а высоконенасыщенные жирные кислоты — около 60% и более (ARA, DHA, ALA, EPA).
III. Взаимосвязь между белками головного мозга и старением
Старение мозга — это не единичный случай, а системный, прогрессирующий процесс, включающий постепенное нарушение баланса белкового гомеостаза мозга, постепенное истощение функциональных белков и постепенное накопление аномальных белков.
3.1 Комплексный спад сети белкового гомеостаза
Все три основные клеточные системы, отвечающие за контроль качества белков, демонстрируют функциональный спад:
3.1.1 Сниженная эффективность синтеза и сворачивания белка
В стареющих клетках снижается скорость и точность синтеза белка, что увеличивает вероятность образования неправильно свернутых белков.
В результате стресса эффективность молекулярных шаперонов снижается, они перестают должным образом помогать вновь синтезированным белкам в правильном сворачивании или восстанавливать поврежденные белки.
3.1.2 Снижение эффективности системы расщепления белка
Убиквитин-протеасомная система демонстрирует пониженную активность, неспособная оперативно удалять поврежденные или неправильно свернутые белки; сборка протеасомы также нарушается окислительным повреждением, что ускоряет снижение ее функциональности.
Аутофагически-лизосомная система, в частности макроаутофагия и аутофагия, опосредованная шаперонами, демонстрирует значительно сниженную эффективность. Это препятствует эффективному удалению аномальных белковых агрегатов и поврежденных органелл (например, митохондрий).
3.1.3 Нарушение транспорта и распределения белков
Нарушение функций структурных белков препятствует эффективной доставке необходимых для синапсов белков и митохондрий к дистальному концу клетки, в то время как продукты жизнедеятельности не могут быть оперативно транспортированы обратно в тело клетки для переработки.
3.2 Накопление и токсичность аномальных белков
Нарушение гомеостаза приводит к постепенному накоплению в головном мозге различных «отходов», непосредственно повреждающих нейроны.
3.2.1 Накопление неправильно свернутых и склонных к агрегации белков
Появляются растворимые олигомеры белков, таких как β-амилоид, тау-белок и α-синуклеин.
3.2.2 Накопление аномально посттрансляционно модифицированных белков
Модифицированные AGEs белки: конечные продукты гликирования накапливаются в стареющем мозге, вызывая образование поперечных связей между белками, их уплотнение, потерю функций и активацию воспалительных процессов.
Гиперфосфорилированный белок Тау: Нормальный белок Тау отсоединяется от микротрубочек, теряя свою функцию стабилизации цитоскелета.
3.3 Потеря ключевых функциональных белков
Помимо увеличения количества «отходов», «полезные» белки также уменьшаются или перестают функционировать.
3.3.1 Снижение уровня белков, связанных с синапсами
Снижение экспрессии ключевых пресинаптических (например, синаптофизина, SNAP-25) и постсинаптических (например, PSD-95, NMDA/AMPA-рецепторов) белков напрямую приводит к атрофии синаптических структур и функциональному ухудшению, формируя непосредственную молекулярную основу нарушений обучения и памяти.
3.3.2 Снижение уровня нейротрофических факторов и их рецепторов
Ослабление синтеза и передачи сигналов нейротрофического фактора головного мозга (BDNF), фактора роста нервов (NGF) и других приводит к недостаточному количеству поддерживающих сигналов для выживания, роста и пластичности нейронов.
3.3.3 Снижение активности белков репарации ДНК и теломеразы
Это приводит к геномной нестабильности и клеточному старению, ухудшая долгосрочное выживание нейронов.
3.4 Системные последствия
Упомянутые выше изменения на белковом уровне запускают каскад реакций, кульминацией которых является старение мозга:
3.4.1 Дисфункция митохондрий: Поврежденные белки накапливаются в митохондриях, снижая выработку АТФ и усугубляя окислительный стресс.
3.4.2 Хроническое нейровоспаление: Глиальные клетки (микроглия и астроциты) постоянно активируются аномальными белками (например, олигомерами Aβ), превращаясь из «защитников» в «разрушителей» путем высвобождения провоспалительных факторов, повреждающих здоровые нейроны.
3.4.3 Потеря синаптической пластичности: Дисбаланс ключевых белков ослабляет долговременную потенциацию, ухудшая формирование и консолидацию памяти.
3.4.4 Снижение эффективности нейронной сети: повреждение миелиновых белков в белом веществе и нарушение аксонального транспорта замедляют передачу информации между областями мозга и ухудшают синхронизацию.
3.4.5 Нарушение регуляции аутофагии и апоптоза: В конечном итоге приводит к избирательной потере нейронов, особенно в критически важных областях, таких как гиппокамп и префронтальная кора.
IV. Механизм действия и эффекты питания, не связанного с дегенеративными изменениями головного мозга.
Питание, не вызывающее дегенерации головного мозга, не только обеспечивает комплексное, сбалансированное поступление питательных веществ, необходимых для роста и восстановления мозговой ткани, включая белки, фосфолипиды, гиалуроновую кислоту, альфа-линоленовую кислоту, эйкозапентаеновую кислоту и т. д., но, что более важно, доставляет их естественные активные структуры. Эти структуры обладают особыми функциями, выходящими за рамки простого питания.
4.1 Ингибирование функциональной деградации белков головного мозга
Механизм действия: Трехмерная структура неденатурированного ультрадисперсного порошка нейротрофических клеток головного мозга устойчива к деградации под действием желудочного сока, что обеспечивает прямой физический контакт с кишечной и лимфатической системами. Это активирует незрелые Т-клетки, которые трансформируются в специфические регуляторные Т-клетки (Treg). Treg-клетки модулируют эффекторные Т-клетки, предотвращая ошибочные иммунные атаки на белки головного мозга, выделяют противовоспалительные факторы для подавления хронического воспаления и замедляют деградацию белков головного мозга.
4.2. Содействие структурному формированию и аномальному восстановлению белков головного мозга.
Механизм действия: Неденатурированный нейро-ультратонкий порошок головного мозга содержит неповрежденные первичные и вторичные ковалентные сшивающие структуры. Кишечные пищеварительные ферменты избирательно расщепляют первичные (пептидные) ковалентные сшивки, сохраняя при этом вторичные ковалентные сшивки. В результате этого процесса образуются малые молекулы, такие как сшитые пептиды и конъюгированные пептиды, или они служат субстратами для специфических ферментов, усиливая каталитическую активность; или функционируют как молекулярные шапероны для восстановления аномальных структур; или индуцируют формирование белковых структур головного мозга и восстановление аномальных структур посредством множественных путей, таких как передача сигналов.
4.3 Обеспечение питательными веществами
Механизм: Малые молекулы, такие как сшитые пептиды и конъюгированные пептиды, могут далее распадаться на отдельные аминокислоты, непосредственно участвуя в синтезе белков головного мозга.
4.4 Эффект
Этот двойной регуляторный эффект, заключающийся в ингибировании деградации и стимулировании синтеза, обеспечивает значительно большую эффективность, чем роль денатурированных белков или пептидов в обеспечении питательными веществами.
V. Тканевые материалы
5.1 Регуляция, зависящая от структуры ткани
Основные функции неденатурированного питания (иммуномодуляция, регуляция активности) достигаются за счет естественных активных структур. Эти структуры обладают высокой специфичностью, оказывая регулирующее воздействие только на белки с идентичной структурой.
5.2 Соответствующее происхождение ткани
Для питания мозга неденатурированными продуктами используется свежая ткань головного мозга крупного рогатого скота или свиней, включая головной мозг, средний мозг и мозжечок. Следовательно, ее белковая структура гомологична структуре белков, обнаруженных в головном мозге, среднем мозге и мозжечке.
VI. Подходящие популяции
6.1 Лица, нуждающиеся в развитии и росте головного мозга: младенцы, дети младшего возраста и лица, нуждающиеся в поддержке развития головного мозга и нервной системы.
6.2. Лица, стремящиеся к омоложению мозга или устранению его дисфункции.
Неденатурированный мышечный белок - здоровье мышц
I. Обзор
Ультратонкий порошок неденатурированного мышечного белка — это неденатурированный функциональный пищевой ингредиент, получаемый из свежей говяжьей вырезки или куриной грудки. Он производится в соответствии с действующими стандартами с использованием сверхнизкотемпературного сверхтонкого измельчения (запатентованная технология) и сверхнизкотемпературной технологии разделения на основе принципа воздушной камеры (собственная технология).
II. Соотношения и функции состава
2.1 Общая классификация

2.2 Белковый состав и функции
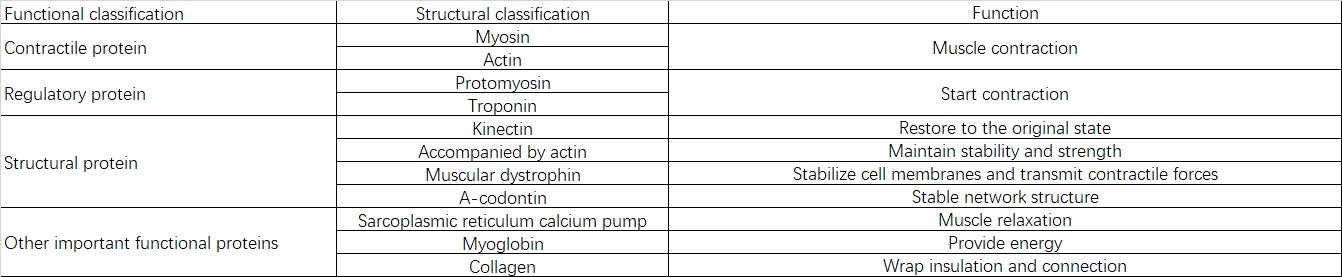
III. Причины саркопении
Саркопения — это прогрессирующая потеря массы, силы и функции скелетных мышц, являющаяся результатом множества взаимосвязанных факторов.
3.1 Дисбаланс между синтезом и деградацией белка (основная патология)
3.1.1 Упрощенный синтез
Скорость синтеза мышечного белка снижается с возрастом. Даже при стимуляции (например, с помощью питания/физических упражнений) синтетический ответ ослабевает, что приводит к резистентности к синтезу белка.
3.1.2 Усиленная деградация
Чрезмерная активация убиквитин-протеасомной системы и аутофагически-лизосомной системы в сочетании с повышением экспрессии таких генов, как Atrogin-1 и MuRF1.
3.2 Нарушение гормонального баланса и сигнальных путей
3.2.1 Снижение уровня анаболических гормонов
Значительное нарушение оси гормон роста/инсулиноподобный фактор роста-1.
Снижение уровня половых гормонов (тестостерона/эстрогена).
3.2.2 Ингибирование анаболических путей:
Инсулинорезистентность: снижение эффективности усвоения и использования аминокислот в мышцах.
Подавление ключевых анаболических путей: например, снижение способности к активации сигнального пути mTOR.
Повышенный уровень миостатина: подавляет рост мышц.
Усиление катаболических сигналов: относительное или абсолютное увеличение уровня глюкокортикоидов способствует расщеплению белков.
3.3 Хроническое воспаление
Старение сопровождается системным, хроническим, вялотекущим воспалением с повышенным уровнем провоспалительных цитокинов (например, ИЛ-6, ФНО-α).
IV. Механизм действия и эффекты
Ультратонкий порошок неденатурированного мышечного протеина не только обеспечивает комплексное, сбалансированное питание, необходимое для роста мышц, но и обладает «активными структурными» свойствами.
4.1 Механизм и эффекты ингибирования деградации
Механизм действия: Микропорошок из неденатурированного мышечного белка содержит множество неденатурированных мышечных белков с неповрежденной трехмерной структурой. Эти белки устойчивы к деградации желудочным соком и достигают тонкой кишки в неизмененном виде. Они фагоцитируются М-клетками, что обеспечивает специфический физический контакт с лимфатической системой кишечника, активируя незрелые Т-клетки для трансформации в специфические регуляторные Т-клетки (Treg). Это модулирует ошибочные атаки эффекторных Т-клеток на различные мышечные белки, секретирует противовоспалительные факторы и подавляет хроническое воспаление.
Эффект: замедляет распад мышечного белка.
4.2 Механизм и эффекты стимулирования синтеза
Механизм действия: Неденатурированный ультрадисперсный порошок мышечного белка содержит неповрежденные первичные и высокоуровневые ковалентные сшивки. Кишечные пищеварительные ферменты могут специфически отбирать и расщеплять первичные (пептидные) ковалентные сшивки, сохраняя при этом высокоуровневые ковалентные сшивки. В результате образуются небольшие молекулы, такие как сшитые пептиды и конъюгированные пептиды, которые затем регулируют каталитическую активность протеаз различными способами — в качестве субстратов, молекулярных шаперонов или сигнальных молекул — для усиления синтеза мышечного белка. Дальнейшее расщепление на отдельные аминокислоты обеспечивает непосредственное участие в синтезе мышечного белка.
Эффект: Способствует синтезу мышечной ткани.
Такое двойное действие — ингибирование деградации и одновременное стимулирование синтеза — обеспечивает значительно большую эффективность, чем денатурированные белки или пептиды.
V. Тканевые материалы
5.1 Структура, определяемая тканями
Основные функции неденатурированного питания (иммуномодуляция, регуляция активности) достигаются за счет естественных активных структур. Эти структуры обладают высокой специфичностью, оказывая регулирующее воздействие только на белки с идентичной структурой.
5.2 Тканеспецифическое происхождение
Неденатурированные мышечные питательные вещества получают из говяжьих сухожилий или куриной грудки — тканей скелетных мышц. Говяжьи сухожилия имеют 95% структурного сходства, а куриная грудка — 85%.
VI. Подходящие популяции
6.1 Спортсмены: Лица, занимающиеся силовыми тренировками и наращиванием мышечной массы.
6.2 Саркопения: подавляет деградацию мышечной ткани и способствует синтезу мышечной ткани.
Неденатурированный белок селезенки — здоровье иммунной системы
I. Обзор
Ультратонкий порошок неденатурированного белка селезенки — это неденатурированный функциональный пищевой ингредиент, производимый из свежей бычьей селезенки. Он изготавливается с использованием технологии сверхнизкотемпературной сверхтонкой обработки (национальный патент на изобретение) и технологии сверхнизкотемпературного разделения на основе принципа воздушной камеры (собственная технология) в соответствии с действующими стандартами.
II. Соотношения компонентов и функции

2.1 Состав и функции основных белковых комплексов иммунного ответа
2.1.1 Иммунорелевантные белки (основной функциональный комплекс): наиболее характерная и функциональная фракция среди белков селезенки.
Состав: Антитела (иммуноглобулины IgG, IgM и др.) и их фрагменты.
Функция: Основные исполнители гуморального иммунитета.
2.1.2 Белки главного комплекса гистосовместимости
Состав: молекулы MHC класса I и MHC класса II, широко экспрессируемые на поверхности антигенпрезентирующих клеток.
Функция: Захват фрагментов белков патогенов (антигенов) и их «презентация» Т-лимфоцитам.
2.1.3 Белки системы комплемента
Состав: Множество компонентов комплемента, включая C3, C4 и активаторы комплемента.
Функция: Формирование «комплементарного мембраноатакующего комплекса» для непосредственного прокалывания мембран патогенов; действие в качестве «хемокинов» для привлечения иммунных клеток к местам инфекции; маркировка патогенов для стимулирования фагоцитоза (комплементарная активность).
2.1.4 Цитокины и хемокины
Состав: интерлейкины, интерфероны, факторы некроза опухолей и др.
Функция: Выступают в качестве «сигнальных молекул» между иммунными клетками, точно направляя инициирование, усиление, подавление или прекращение иммунных реакций и координируя поведение различных типов иммунных клеток.
2.1.5 Рецепторы распознавания образов
Состав: Толл-подобные рецепторы и др.
Функция: Распознавание консервативных молекулярных паттернов патогенов для инициирования быстрых реакций врожденного иммунитета.
2.2 Белки, участвующие в обработке эритроцитов
2.2.1 Ферменты, расщепляющие гемоглобин
Состав: гемоксигеназа, биливердинредуктаза и др.
Функция: Расщепление гемоглобина в стареющих эритроцитах с высвобождением ионов железа для повторного использования и превращением гема в билирубин.
2.2.2 Белки, участвующие в хранении и транспортировке железа
Состав: Ферритин является наиболее важным представителем этого комплекса.
Функция: Безопасно и эффективно хранит и нейтрализует железо, предотвращая образование вредных свободных радикалов из свободного железа, и высвобождает железо по мере необходимости для организма.
2.3 Структурные и каркасные белки
Состав: белки внеклеточного матрикса (например, коллаген, фибронектин) и молекулы клеточной адгезии.
Функция: Формирование физического каркаса белой пульпы селезенки (периартериальные лимфатические оболочки, лимфоидные фолликулы) и красной пульпы; обеспечение миграции, удержания лимфоцитов и взаимодействия между иммунными клетками.
2.4 Метаболизм и антиоксидантные ферменты
2.4.1 Антиоксидантные ферменты
Состав: супероксиддисмутаза, каталаза, глутатионпероксидаза.
Функция: нейтрализация активных форм кислорода (АФК) для защиты тканей селезенки от окислительного повреждения.
2.4.2 Ферменты метаболизма нуклеотидов
Состав: пуриннуклеозидфосфорилаза и др.
Функция: Участвует в метаболизме пуринов в лимфоцитах, что имеет решающее значение для функционирования иммунных клеток.
2.5 Белки, участвующие в передаче сигналов и регуляции
Состав: факторы транскрипции, включая NF-κB, белки семейства STAT и др.
Функция: Принимать сигналы, такие как цитокины, перемещаться в ядро для регуляции экспрессии определенных генов и определять судьбу активации, пролиферации или дифференцировки иммунных клеток.
III. Механизм действия и эффекты
Ультрадисперсный порошок неденатурированного белка селезенки содержит неповрежденные, естественно сохранившие свою конформацию белки селезенки. Его основной механизм действия не связан с обеспечением питательными веществами, а скорее представляет собой сложный природный резервуар биоактивных молекул. Он поставляет регуляторные сигналы и функциональные модули в организм, в частности, в иммунную систему, оказывая тем самым эффект «питательного иммунитета».
3.1 Местное воздействие на кишечный иммунитет
3.1.1 Пассивные иммунные эффекты
Неденатурированные ультрадисперсные частицы белка селезенки содержат большое количество иммуноглобулинов, которые достигают кишечника в неизмененном виде, сохраняя структурную защиту. Их антиген-связывающие фрагменты (Fab-области) сохраняют активность в течение определенного периода, обеспечивая местный пассивный иммунитет. Эти Fab-фрагменты могут: - Непосредственно нейтрализовать определенные вирусы, бактерии и их токсины, делая их неспособными к инфекции или токсическому воздействию; - Прикрепляться к патогенам, блокируя их проникновение; - Образовывать комплексы с патогенами или антигенами, которые стимулируют перистальтику кишечника и способствуют инкапсуляции в слизь, ускоряя их выведение с калом и сокращая время их задержки и вредное воздействие в кишечнике.
3.1.2 Поддержание барьерной функции кишечника
Снижение воспалительного повреждения: быстро устраняя патогены и токсины, препарат уменьшает прямое воздействие на эпителиальные клетки кишечника и связанную с этим чрезмерную воспалительную реакцию, тем самым защищая целостность кишечного барьера.
Поддержка восстановления: Некоторые исследования показывают, что другие компоненты в препаратах иммуноглобулинов (например, факторы роста) или сами антитела могут косвенно способствовать восстановлению слизистой оболочки.
3.1.3 Модуляция местной иммунной среды
Буферизация антигенов: В кишечнике — самом большом иммунном органе организма — избыток экзогенных антигенов (включая пищевые и патогенные антигены) может вызывать аномальные иммунные реакции. Пероральный иммуноглобулин связывает и удаляет часть этих антигенов, снижая нагрузку на кишечную иммунную систему и риск ошибочных атак, тем самым помогая поддерживать баланс иммунной толерантности.
Противовоспалительный эффект: за счет снижения стимуляции патогенами посредством вышеупомянутых механизмов, препарат подавляет выработку провоспалительных цитокинов, создавая локальную среду, более благоприятную для восстановления тканей и поддержания микробного баланса.
3.2 Системные иммуномодулирующие эффекты
3.2.1 Индукция периферической иммунной толерантности в виде антигенов
Селезенка содержит белки и антигены, связанные с иммунной системой, в их нативной, неденатурированной форме. Эти вещества могут распознаваться кишечной иммунной системой как «дружественные и знакомые иммунные стимулы», повышая иммунную бдительность и реактивность без запуска интенсивного воспаления (путь оральной толерантности).
3.2.2 Непосредственное предоставление иммунологически активных компонентов и сигнальных молекул
Высвобождает в кишечнике низкомолекулярные пептиды с иммуномодулирующей активностью. Эти пептидные фрагменты функционируют как сигнальные молекулы, распознаваемые иммунными клетками кишечника (например, клетками пейеровых бляшек), мягко стимулируя и тренируя иммунную систему — подобно «иммунной репетиции».
3.2.3 Взаимодействие с хранилищем иммунной информации
Возможность индукции оральной толерантности: Согласно теориям, при определенных аутоиммунных заболеваниях употребление в пищу тканей, содержащих собственные антигены, может способствовать индукции иммунной толерантности.
IV. Тканевые материалы
4.1 Зависимость от структуры ткани
Основные функции неденатурированного питания (иммуномодуляция, регуляция активности) достигаются за счет естественных активных структур. Эти структуры обладают высокой специфичностью, оказывая регулирующее воздействие только на белки с идентичной структурой.
4.2 Соответствующее происхождение ткани
Селезенка, богатая разнообразными иммунными клетками и факторами, по своему происхождению соответствует иммунной ткани.
V. Подходящие популяции
5.1 Лица с ослабленным иммунитетом
5.2 Лица с расстройствами пищеварения
Неденатурированный эластин — здоровье сосудов
I. Обзор
Ультрадисперсный порошок неденатурированного эластина — это неденатурированный функциональный пищевой ингредиент, получаемый из свежих затылочных связок или аорт крупного рогатого скота. Он производится в соответствии с соответствующими стандартами с использованием ультрамикронизации при сверхнизких температурах (запатентованная технология национального изобретения) и технологии сепарации при сверхнизких температурах на основе принципа «воздушной камеры» (собственная технология).
II. Соотношения компонентов
2.1 Состав ультрадисперсного порошка неденатурированного эластина (затылочная связка крупного рогатого скота) и его лиофилизированного порошка

2.2 Состав ультрадисперсного порошка неденатурированного эластина (аорта крупного рогатого скота) и его лиофилизированного порошка

III. Естественная активная структура
3.1 Первичная структура
Аминокислотная последовательность эластина состоит из чередующихся гидрофобных сегментов (богатых валином, аланином, глицином и пролином) и сшитых сегментов (богатых лизином и аланином), расположенных тандемно.
3.2 Высшая структура
Неупорядоченная аминокислотная последовательность белка эластина препятствует образованию регулярных вторичных структур, в которых отсутствуют традиционные структуры II, III или IV уровней. Вместо этого он образует трехмерную неупорядоченную сетчатую полимерную структуру посредством ковалентных поперечных связей, таких как поперечные связи эластина.
IV. Механизм действия и эффекты
Эластин — это непитательный белок, основная функция которого заключается не в обеспечении аминокислотами, а в наличии уникальной «естественной активной структуры», оказывающей структурное воздействие.
4.1 Механизм и эффекты периферической иммунной регуляции
4.1.1 Механизм: Многомерная структура, уникальная для неденатурированного эластина, активирует дифференцировку лимфоидных Т-клеток пейеровых бляшек в специфические регуляторные Т-клетки (Treg). Это модулирует ошибочную атаку иммунной системы на эндогенный эластин и подавляет воспалительные реакции.
4.1.2. Эффекты: Замедляет деградацию эластина. С возрастом (начиная примерно с 25-30 лет) способность организма синтезировать эластин резко снижается. Одновременно с этим, из-за нарушения иммунной регуляции, существующие эластиновые волокна разрушаются, затвердевают и становятся дисфункциональными вследствие гликирования, окислительного стресса (повреждения свободными радикалами) и ферментативной деградации (например, матриксными металлопротеиназами). Это является основной причиной уплотнений сосудов в организме человека.
4.2 В качестве клеточной сигнальной молекулы (наиболее заметная функция)
4.2.1 Механизм: Поверхность неповрежденных молекул эластина содержит специфические биоактивные домены (например, домены связывания с клетками, хемотаксические домены). Сохраняя свою нативную конформацию, эти домены могут специфически связываться с рецепторами (например, белками, связывающими эластин) на поверхности фибробластов, эндотелиальных клеток и других клеток. Это связывание сигнализирует клеткам о необходимости стимулировать собственный синтез дополнительного коллагена, эластина и гиалуроновой кислоты. Этот процесс индуцирует эндогенную регенерацию, которая гораздо важнее, чем простое пополнение запасов сырья.
4.2.2 Эффект: Способствует существенному улучшению эластичности и упругости, восстанавливая эластичность таких тканей, как кровеносные сосуды.
4.2 В качестве субстрата, модулирующего активность фермента
4.2.1 Механизм: Неденатурированный эластин может расщепляться в кишечнике на сшитые пептиды, содержащие первичные и вторичные ковалентные сшивающие структуры. Эти сшитые пептиды служат субстратами для соответствующих ферментов, модифицирующих структуру, тем самым повышая их каталитическую активность.
4.2.2 Действие: Способствует формированию или восстановлению эластиновых структур.
4.3 В качестве структурного шаблона и каркаса
4.3.1 Механизм: Неденатурированный эластин, сохраняющий крупные структурные фрагменты и неденатурированные сшитые структуры, может функционировать как «направляющий каркас» in vitro или in vivo. Он направляет вновь синтезированные молекулы-предшественники эластина к упорядоченному расположению и правильному сшиванию, способствуя формированию функциональных новых сетей эластичных волокон.
4.3.2 Действие: Способствует восстановлению поврежденных, деформированных эластичных волокон, вызванных фотостарением или старением.
V. Тканевые материалы
5.1 Зависимость от структуры ткани
Функции неденатурированных белков (иммуномодуляция, регуляция активности) зависят от их структуры. Их структура обладает высокой специфичностью, регулируя только белки с идентичной структурой.
5.2 Соответствующие источники тканей
Ткани, богатые эластином, такие как кровеносные сосуды, кожа и альвеолы.
VI. Подходящие кандидаты
6.1 Состояние сосудистой системы: гипертония, гиперлипидемия, атеросклероз.
6.2. Борьба с морщинами: дряблость кожи, морщины, потеря эластичности.
6.3 Легочная недостаточность: пациенты с эмфиземой, хронической обструктивной болезнью легких (ХОБЛ) и др.





